4. Гены
Вернемся снова к опытам Менделя: наблюдается известный параллелизм между гипотетическими наследственными факторами Менделя и реально наблюдаемыми хромосомами. Подобно хромосомам, эти факторы передаются поодиночке через зрелые зародышевые клетки; во время оплодотворения они располагаются попарно; они разделяются или расщепляются вновь, когда последующая генерация образует свои зародышевые клетки. Имеется лишь одно препятствие расценивать хромосомы как менделевские факторы - их численность слишком мала. Например, горох имеет только 7 пар хромосом. Мендель изучал семь пар альтернативных признаков, и нет сомнения, что число их он мог бы увеличить. Для объяснения наследственной конституции целого организма гороха, мыши, мухи или человека требуются десятки тысяч наследственных факторов. Выход из этой дилеммы состоит в том, чтобы рассматривать хромосомы не как наследственные факторы, а как переносчики, на которых эти факторы передаются от потомства к потомству. Истинность этого предположения подтверждена многочисленными экспериментами, и теперь уже больше нет никакого сомнения, что наследственные факторы, или гены, располагаются вдоль хромосом, и каждая хромосома несет сотни или даже тысячи генов.
Гены слишком малы, чтобы разглядеть каждый отдельно, но на определенных стадиях вдоль многих хромосом обнаруживаются узелки, утолщения или диски, которые указывают на подразделение хромосом на отдельные гены. В особенности хорошо это видно на разных стадиях мейоза. На рис. 4, а, где показана пара хромосом ржи в начале мейоза, можно заметить участки темноокрашенных узелков и слабоокрашенные связующие их нити. У двух хромосом-партнеров каждый отрезок сходен, спаривание является очень точным. Каждый узелок одной хромосомы ложится рядом с соответствующим узелком ее партнера. По всей вероятности, эти узелки - гены, а не хромосомы в целом направляют процесс спаривания. Такая способность генов находить и притягивать к себе своих двойников является замечательным свойством, которому долгое время наука не могла дать удовлетворительного объяснения. Позже, во время мейоза, эта структура исчезает, так как хромосомы свиваются и превращаются в компактные образования (см. рис. 2, з). Но в дальнейшем, когда в следующем поколении будут развиваться половые клетки, каждая хромосома вновь приобретает характерное строение. Опытный цитогенетик и цитолог могут узнать каждую из семи хромосом ржи по расположению ее узелков.
В слюнных железах личинок некоторых мух обнаружены гигантские хромосомы с красивыми и сложными участками темных и светлых узелков-дисков. На рис. 4, б показаны такие хромосомы в слюнных железах дрозофилы. Они в сотни раз больше, чем обычные хромосомы, и каждая из них имеет набор характерных участков с дисками. Эти хромосомы также во много раз толще обычных хромосом, потому что каждая состоит из пучков с огромным числом нитей с совершенно одинаковыми дисками. Очень интересен путь возникновения этих пучков, потому что он показывает чрезвычайную точность процесса удвоения хромосом. У молодой личинки хромосомы в слюнных железах еще очень тонки. По мере роста личинки увеличиваются также размеры слюнной железы, но растет она своеобразно. Клетки вместо увеличения их числа путем деления начинают расти все больше и больше, без деления. Увеличивается в размерах также их ядро. Внутри ядра хромосомы начинают удваиваться, подобно тому, как это происходит перед каждым делением клетки, а так как мейоза не наступает, новые хромосомы не отделяются от старых. В результате новые и старые хромосомы остаются рядом друг с другом и каждая новая вновь начинает удваиваться. Все это происходит многократно с большой точностью, так что каждый пучок образует совершенно подобный себе пучок и вся хромосома выглядит как единая толстая, состоящая из дисков нить. Кроме этого, хромосомы-партнеры в слюнных железах так тесно спарены в мейозе, и то, что мы видим в микроскоп и считаем одной хромосомой, на самом деле представляет собой пару, а ядра слюнных желез дрозофилы несут четыре хромосомы вместо восьми, которые наблюдаются в обычных клетках (см. рис. 2, в). Схема дисков отражает схему расположения генов. Это можно часто наблюдать в случаях, когда хромосома теряет маленький кусочек под влиянием радиации или другого повреждающего воздействия. Процесс репликации (создания копии) хромосом идет столь точно, что все потомки пораженной хромосомы будут получаться без такого исчезнувшего кусочка, и когда пораженная хромосома попадает в гамету, например яйцеклетку, то развившиеся особи получают хромосому с нехваткой исчезнувшего кусочка во всех клетках. Рис. 5 и 6 показывают, что в таких случаях происходит.

Рис. 4. Хромосомы, зародыши и уродство различных организмов: а - пара хромосом ржи в начале мейоза; б - гигантские хромосомы в слюнных железах дрозофилы; в - эмбрионы бесхвостой (слева) и нормальной мыши в одном возрасте; г - водяночный теленок; д - 'ампутированный' теленок
Левая часть рис. 5 показывает нормальное крыло дрозофилы, а над ним, под гораздо большим увеличением, маленький участок одной из нормальных хромосом слюнной железы. Стрелка указывает на диск, в котором находится ген, управляющий образованием формы крыла. Правая часть рис. 5 показывает этот же участок хромосомы у мухи, которая получила поврежденную хромосому от отца или матери, облученных рентгеновскими лучами. Если вы вспомните, что хромосомы слюнных желез состоят из двух пучков хромосом, образованных из двух хромосом-партнеров, станет ясно, почему на месте стрелки отсутствует лишь половина диска. Тем не менее нехватки одного из двух генов-партнеров было достаточно для нарушения нормального развития крыльев и появления уродства (см. рис. 5, справа). Насекомые с утерей диска в обеих хромосомах-партнерах столь сильно повреждены, что умирают на стадии раннего эмбрионального развития. Подобный случай показан на рис. 6. Левая часть рисунка изображает нормально выросшую зеленую кукурузу, а под ним при значительно более сильном увеличении - кончики одной определенной хромосомной пары. На кончике каждой из хромосом-партнеров видны довольно крупные узелки. В этом узелке среди генов находится ген, присутствие которого совершенно обязательно для того, чтобы в клетках образовывалось зеленое вещество - хлорофилл, необходимый для жизни растения. Правая часть рисунка показывает,- что происходит, если этот крупный узелок исчезает из обеих хромосом: растение отмирает очень рано, имея вид желтого и больного проростка. Если обратиться еще раз к левой половине рис. 6, можно уловить отличие от того, что наблюдалось с геном крыла дрозофилы. Там этот ген не мог обеспечить развития нормального крыла без помощи гена-партнера; хлорофилловый ген вполне дееспособен, даже если он действует в одиночку; растения совершенно нормальные, если одна хромосома нормальна, а другая лишена узелка. Дело здесь не в различиях генов кукурузы и дрозофилы и не в генах, управляющих формой крыла или образованием хлорофилла. Дело здесь в эффективности, с которой гены выполняют свои функции. Некоторые гены эффективны в одиночку, другие для проявления эффекта требуют кооперации со своими партнерами.
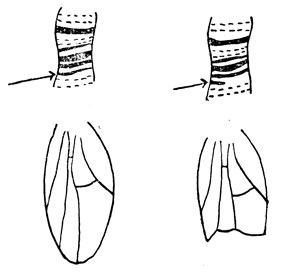
Рис. 5. Нехватка одного из генов крыла дрозофилы
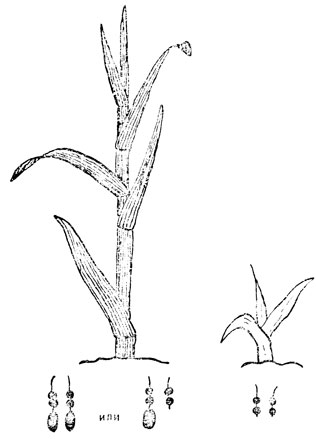
Рис. 6. Нехватка хлорофиллового гена кукурузы
Каждый ген играет роль в определенном процессе развития, например: в образовании хлорофилла, развитии окраски или размеров или у высших животных в развитии психических способностей. Гены-партнеры управляют тем же процессом, но действуют иным путем; два гена-партнера у гороха управляют формой семян: один определяет, что семена получаются гладкими, а другой делает их морщинистыми. Гены-партнеры, дающие различный результат, называются аллеломорфами, или аллелями. Слово "аллеломорф" означает разнеформенный; согласно новейшим представлениям, это можно понимать как изменение химического строения в основном того же самого гена.
Все процессы развития чрезвычайно сложны, а потому не удивительно, что очень много различных пар генов принимает участие в развитии, например, глаза, мозга или цветка (см. гл. 23). Даже такой, казалось бы, простой процесс, вроде образования витамина в бактериальной клетке, проходит через ряд ступеней и управляется несколькими генами. У бактерий гены, управляющие одним и тем же процессом, имеют тенденции располагаться по соседству, как будто это облегчает их совместные действия, но у других организмов этого не наблюдается. Гены, связанные с самыми разнообразными процессами, могут лежать совершенно рядышком в той же хромосоме, тогда как гены, регулирующие связанные процессы, могут быть расположены поодаль. На рис. 7 показано расположение нескольких из известных генов на одной хромосоме дрозофилы. Помечены лишь гены, связанные с развитием глаз и крыльев; помимо этих генов имеется еще много генов, управляющих развитием этих органов, которые лежат на данной хромосомной паре и на трех других. Можно видеть, как по-разному действуют представленные на схеме гены: крыловой ген wl управляет формой крыла, тогда как ген w2 управляет развитием правильного жилкования крыла. Для других генов такое разделение не доказано: так, все глазные гены el, e2, е3 и е4 управляют окраской глаз. Однако из исследований по развитию особи известно, что каждый из них играет свою собственную роль в этом процессе.
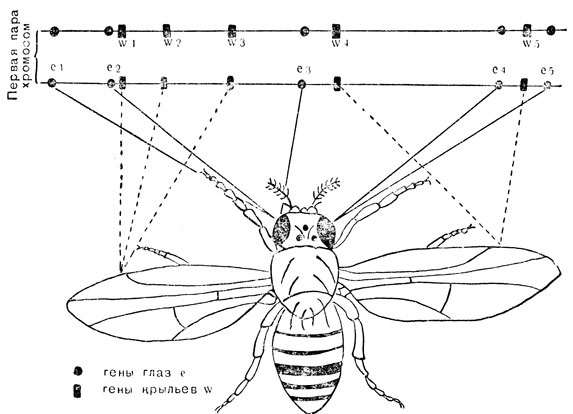
Рис. 7. Положение некоторых генов крыла и глаза на одной из хромосом у дрозофилы
Прежде чем перейти к следующей главе, полезно подвести итог тому, что мы узнали о генах. Гены, подобно хромосомам, расположены попарно, с хромосомами они расщепляются в разные гаметы при мейозе и опять сходятся в процессе оплодотворения, причем один член каждой пары приобретается от отца, а другой - от матери. Для каждого вида организмов строго фиксировано распределение генов на хромосомах, и гены-партнеры занимают те же самые места на хромосомах-партнерах. Аллеломорфы представляют собой гены-партнеры, занимающие то же место на той же хромосомной паре, и управляют тем же процессом во время развития, но другим путем.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'