12. Как наследуются количественные признаки?
Количественные признаки - это все мерные признаки: рост (в сантиметрах), удой молока (в литрах), жирность молока (в процентах) и т. п. Анализ наследования количественных признаков значительно затруднен. Первое, с чем сталкивается генетик, - это непрерывный характер изменчивости.
Уже при анализе наследования окраски зерна у пшеницы, обусловленной полимерными генами, Н. Г. Нильсон-Эле установил непрерывный ряд изменчивости окраски зерна растений в F2 от белой до интенсивно-красной. Число гетерозиготных локусов, отвечающих за данный признак, можно было установить по количеству появившихся рецессивных потомков: 1/16 белозерных - расщепление идет по двум локусам, 1/64 - по трем, 1/256 - по четырем, 1/1024 - по пяти и т. д., т. е. 1/4n. Таким образом можно выявить только число локусов, по которым родители различаются.
Число пар факторов n, отвечающих за исследуемый признак, вычисляется по формуле
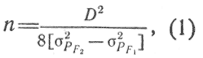
где D - разница между средними арифметическими признаков у родителей; σPF1, σPF2 - фенотипическая изменчивость F1 и F2 соответственно.
Наследование количественных признаков подчиняется менделевским законам расщепления: чем больше пар аллелей влияет на признак, тем больше варьирование по данному признаку. Гибриды в F1 в отношении количественных признаков оказываются промежуточными между родительскими формами, в F2 наблюдается варьирование признака от наименьшего значения одного родителя до наибольшего значения другого.
В F2 количество генотипов равно 3n, но генетических значений меньше, так как они зависят только от количества доминантных аллелей, а не от их локализации. Например, если признак контролируется двумя парами факторов, в F2 будет 32 = 9 генотипов. Однако формы А1А1а2а2, a1a1A2A2 и А1а1А2а2 имеют одинаковое генетическое значение, так как включают по две доминанты. Это касается и форм с одной и тремя доминантами. В нашем примере генетических значений пять: 4А, 3A, 2А, 1А, 0А, т. е. 2n + 1.
Распределение потомства соответствует биномиальной кривой, а коэффициенты генетических значений - показателям степени членов разложения бинома Ньютона:
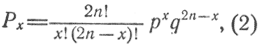
где Рх - вероятность числа особей определенного генетического значения; 2n - число аллелей, контролирующих исследуемый признак; х - число доминантных аллелей определенного генетического значения; р - вероятность доминантного аллеля в популяции; q - вероятность рецессивного аллеля в популяции; ! - факториал (произведение натурального ряда чисел от большего до самого малого, например 5! равно 5·4·3·2·1).
Если х = 0 или х = 2n, формула (2) упрощается:
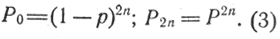
Практически вычисления можно сократить: частота особей определенного генетического значения равна частному от деления произведения частоты предыдущего члена и его генетического значения на число предшествующих членов:
число доминантных генов в генотипе: 4А 3А 2А 1А 0А частота генотипов: 1/16 4/16 6/16 4/16 1/16
Имеются и другие способы подсчета расщепления по количественным признакам.
Кроме генетических свойств организма существует ряд внешних по отношению к нему воздействий, которые могут сглаживать наследственные различия и еще более способствовать непрерывному характеру изменчивости этих признаков. Общая фенотипическая изменчивость σР2 слагается из двух факторов: генотипической изменчивости σG2 и изменчивости под влиянием окружающей среды σE2:
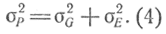
Значение σG2 найдем следующим образом. Изменчивость в F1 определяется лишь условиями окружающей среды, так как особи в F1 по генотипу однородны, т. е. σ2F1 = σ2E; изменчивость в F2 определяется двумя факторами, т. е. σ2F2 = σ2G + σ2Е. Подставляя вместо σ2Е его значение в формулу (4), найдем
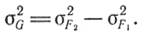
Величину σ2Е можно найти с помощью метода идентичных однояйцевых близнецов. Все наблюдаемые у них различия определяются влиянием условий окружающей среды, т. е. σ2Р = σ2Е.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'