109. Что такое цистрон?
Согласно классическим представлениям ген обозначает единицу генетического материала, которая передается от родителей потомству и может быть обнаружена по ее способности мутировать в различные состояния, рекомбинировать с такими же единицами и функционировать, т. е. определять тот или иной фенотип организма. Такие представления о гене просуществовали вплоть до середины 50-х гг. XX в., до тех пор, пока не была изучена структура гена. Эксперименты С. Бензера показали, что классического гена не существует, поскольку единицей мутации и рекомбинации может служить одна пара нуклеотидов в ДНК. Оставался открытым вопрос о единице функции, и необходимо было выяснить, изменяет ли мутация гена его функцию как единого целого. Для выяснения этого вопроса надо было найти метод, позволяющий определять мутации в различных функциональных единицах.
Чтобы определить, принадлежат ли обе мутации к одному или разным генам, С. Бензер применил цис-транс-тест, или тест на комплементарность (рис. 25). Тест на комплементарность основывается на том, что при заражении штамма Е. coli К двумя бактериофагами - мутантным (неспособным расти на газоне штамма К) и диким фагом r+ - оба штамма развиваются нормально. Значит, нормальный ген дикого типа способен обеспечить функцию, необходимую для размножения на штамме К не только дикого фага, но и rII-мутанта.
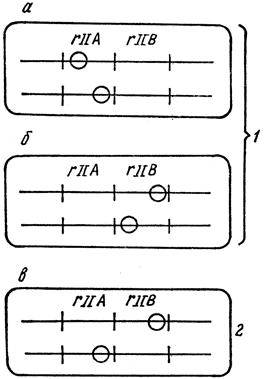
Рис. 25. Тест на комплементарность у бактериофага Т4: а, б, в - мутации в цистронах А, В и в разных цистронах соответственно; 1 - размножение не происходит; 2 - нормальное размножение
Было замечено, что в некоторых случаях не только дикий штамм фага способствует росту мутанта на Е. coli К, но и два мутанта могут помогать друг другу расти на этом штамме, на котором ни один из них в одиночку не способен размножаться. Для выделения таких комплементарных пар мутантов клетки штамма К заражаются одновременно двумя rII-мутантами, чтобы выяснить, смогут ли они при совместном посеве помогать друг другу и давать потомство. Если два мутанта способны к такому совместному размножению, то можно считать, что обе мутации комплементарны и локализованы в различных функциональных участках фагового генома. Если же два мутанта некомплементарны, значит, они затронули одну и ту же функциональную часть генома. Неспособность одного мутанта расти на штамме К свидетельствует о том, что он не может осуществлять какую-то определенную функцию или синтезировать какой-либо белок, необходимый для размножения фага в клетке. В отличие от первого у второго комплементарного мутанта мутация затрагивает другую функцию. Значит, два таких мутанта могут размножаться при совместном посеве. При совмещении двух мутантных геномов в одной бактериальной клетке в ней будут осуществляться все реакции, необходимые для размножения, так как функции, осуществляемые одним мутантом, будут использованы другим.
С помощью теста на комплементарность С. Бензер показал, что мутанты rII разделяются на два класса А и В. Все мутанты одного класса комплементарны любому мутанту другого класса и некомплементарны мутантам того класса, к которому они сами принадлежат. Наличие двух классов мутантов свидетельствует о существовании двух функциональных единиц в пределах области rII генома фага Т4. Каждая функциональная единица связана, по-видимому, с синтезом специфического полипептида, необходимого для роста на штамме К. Генетическую единицу функции, выявленную с помощью цис-транс-теста, С. Бензер назвал цистроном. Термины "ген" и "цистрон" следует употреблять как синонимы.
Путем рекомбинационного анализа можно вычислить длину цистронов А и В в области rII фага Т4. Оказалось, что расстояние между наиболее удаленными точками цистрона А равно примерно пяти единицам карты, а соответствующее расстояние для цистрона В - трем единицам карты. Таким образом, общая длина области rII равна 8% рекомбинации. Исходя из общей длины карты фага Т4 (общая длина карты равна 1500 единицам рекомбинации, 2·105 парам нуклеотидов), можно рассчитать, что цистрон А составляет (5/1500)·2·105 ≈ 700, а цистрон В - (3/1500)·2·105 = 400 пар нуклеотидов.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'