Первая карта хромосомы фага
Херши и Ротман знали, что наследственное вещество фагов, так же как и у всего живого на земле, состоит из генов. В гене закодирована информация для того или иного признака: а значит, и для признака быстроты образования колонии (r), и для круга хозяев (h). У мутантных фагов - мутантные гены. Если изобразить хромосомы двух мутантных фагов- родителей - участников опыта Херши и Ротмана, то они будут выглядеть примерно так.
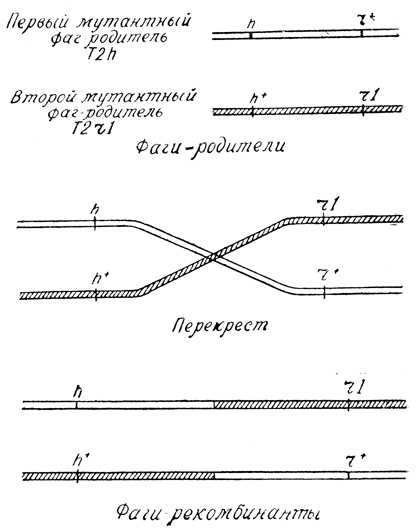
Как видите, у первого фага-родителя мутировал ген, определяющий хозяина (h), у второго - ген скорости размножения (r1). Классический генетик назвал бы пары h и h+ (или r1 и r+) аллелями.
Хромосомы обоих мутантных фагов - образования гибкие и хрупкие. Когда они оказываются рядом, между ними может произойти перекрест (кроссинговер). И если в точке перекреста хромосомы фага разорвутся, а затем воссоединятся, обменявшись местами, то вместо родительских фагов с генами hr+ и h+r1 образуются фаги-рекомбинанты с генами hr1 (мутантный тип) и h+r+ (дикий тип).
Кроссинговер случается в любой точке между генами h и г. Но мы знаем, чем ближе на хромосоме друг к другу гены, тем вероятность кроссинговера между ними меньше, а значит, и количество рекомбинантов в потомстве будет меньше. Следовательно, чем меньше процент таких фагов, тем ближе друг к другу стоят изучаемые гены на хромосоме, и наоборот.
Если теперь вернуться к опытам скрещивания мутанта h с разными r-мутантами, то в итоге их у нас в руках оказались бы такие данные: после скрещивания фага hr+ с мутантом h+rl3 фаги-рекомбинанты образовывались в 1,7 процента случаев, с мутантом r7 - в 12,3 процента, с мутантом r1 - в 24 процентах. Нарисовав хромосому фага и приняв масштаб ее за 1 процент, без труда можно расположить на ней мутантные гены в их относительном положении. Разделим хромосому на 24 единицы (максимальный процент рекомбинантов по этому признаку в нашем случае), тогда ген h и ген r1 будут по краям, r7 - где-то посередине, r13 - около локуса h. Вот и готов один из вариантов хромосомной карты фага Т2.
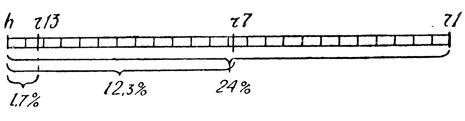
Как будто бы просто, но так только кажется. На деле же составление хромосомных карт-труд поистине титанический. Ведь мы разобрали примерно один из вариантов карты. А всего их было четыре. И ни один не оказался верным. Между тем каждый вариант отнял много сил и энергии у исследователей, и проверяли его не путем рассуждений, а бесконечными экспериментами... Все же труд этот увенчался успехом, и новые скрещивания мутантов фагов Т2 и Т4 позволили, наконец, начертить для них необыкновенно точные карты хромосомы.
Не надо, однако, думать, что кроссинговер происходит между ДНК только родительских частиц фага. Во время размножения он в равной степени "захватывает" также их потомков. Это доказали в опытах по скрещиванию фагов, мутантных по трем генам. Но тройные мутанты помогли понять и другое: хромосома фага представляет собой кольцо. Эта сенсация была бы ошеломляющей для ученых всего мира, если бы Жакоб и Вольман не подготовились к ней заранее, доказав, что хромосома бактерии - тоже кольцо. А если у хозяина фага хромосома кольцевая, то отчего бы ей не быть такой же и у самого фага? Теория кольцевой хромосомы, высказанная учеными Страйзингером и Гарером в 1961 году, была поддержана географами хромосомы фага и позднее получила блестящее подтверждение благодаря прекрасным электронным микрофотографиям кольцевой ДНК фага Т2 (фото 17).
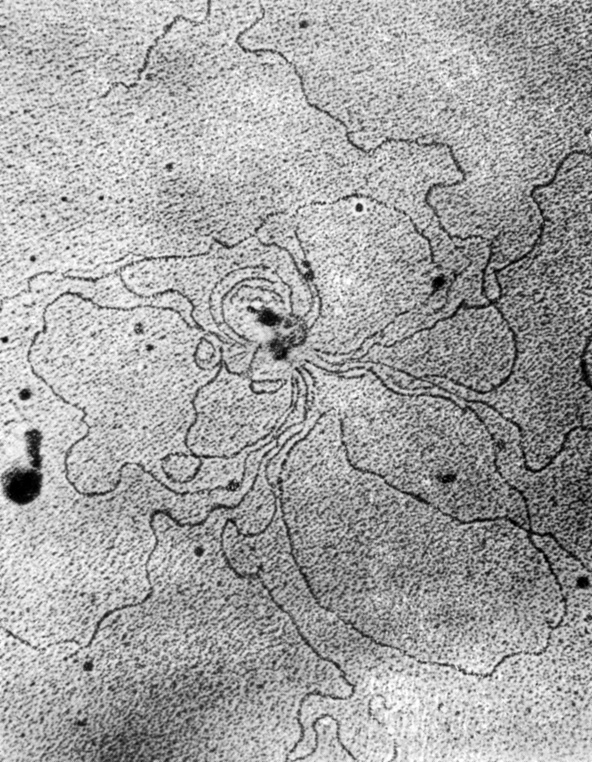
Фото 17. Это ДНК фага Т2, которую сфотографировал в электронном микроскопе А. А. Маныкин. Увеличение в 120 000 раз (лаборатория электронной микроскопии ИЭМ АМН СССР имени Гамалеи)
Фаговые частицы разрушали в тонком слое белка на поверхности раздела вода - воздух. Нить ДНК выплывала из белковой оболочки и располагалась рядом с ней, неповрежденная. И тогда-то исследователи убедились, что она подобна кольцу и не имеет ни конца ни начала.
В составлении карты круглой хромосомы фага принимал участие целый легион мутантов. Среди них были мутанты по признаку чувствительности к температуре, "янтарные" мутанты (не способные размножаться на обычной кишечной палочке), лизоцимные мутанты и многие другие. С каждым годом на обруче хромосомы оставалось все меньше белых пятен. Когда же конец?
Пожалуй, ни один из генетиков не думал столько над этим вопросом, как американский физик Бензер. Ему хотелось раскопать самую сущность гена, перевести единицы хромосомных карт фага, во многом еще абстрактных, на язык химических формул.
Не будет преувеличением сказать, что вся история генетики это в значительной степени история попыток разобраться в сущности гена. Вначале геном называли единицу наследственного материала, которая подчиняется законам Менделя и от которой зависит проявление данного признака.
Постепенно выкристаллизовалось понятие о некоей единице наследственного вещества, передающейся от родителей к потомкам, которую можно выявить благодаря ее способности, во-первых, мутировать (изменяться), во-вторых, рекомбинироваться с другими подобными ей единицами (давать гибриды) и, в-третьих, функционировать (работать, отвечать за развитие определенного признака). Таким образом, ген представлялся ученым как неделимая единица мутации, рекомбинации и строго определенной функции.
Так думали "классические" генетики. Так думал и американский физик Бензер, когда в 1955 году начинал свои опыты с r-мутантами фага Т4. Впрочем, не стоит утверждать это так уж категорически. Будет вернее сказать, что Бензер поставил знак вопроса над "классической" концепцией гена, ибо давно уже в науке зародились сомнения в ее непогрешимости.
Опыты Бензера явились логическим продолжением более ранних работ на дрозофиле, начатых свыше 30 лет тому назад русскими генетиками. В частности, академик Н. П. Дубинин занимался изучением гена, который определял у дрозофилы число и размещение щетинок на спине. У этого гена известно несколько аллелей - измененных форм. Вот почему в лаборатории ученого оказалась целая коллекция дрозофил с разнообразным набором щетинок. Самое удивительное было в том, что, скрещивая некоторые пары мушек, у которых недоставало разных щетинок, Дубинин получал потомство, обладавшее... их полным набором. У мушек-гибридов не развивались только те виды, которых не было у обоих родителей. Исследователь сделал вывод, что, видимо, ген, контролирующий развитие щетинок, состоит из нескольких расположенных рядом друг с другом функционально независимых единиц.
Кроссинговер мог разделять этот ген на части! Но в начале сороковых годов предположения Дубинина потонули в потоке информации и не смогли поколебать почву под ногами классической концепции гена.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'