1.3.1. Гаплоидизации и потеря хромосом
При вегетативных делениях не происходит конъюгация хромосом и, следовательно, обмен частями гомологичных хромосом не осуществляется или осуществляется крайне редко (в этом случае - независимо от потери отдельных хромосом). Потеря - тоже обычно редкое событие, ее частота может быть значительно увеличена при некоторых экспериментальных воздействиях или при введении в генотип специальных мутаций. При потере хромосом (в отличие от мейоза) гены одной хромосомы обнаруживают абсолютное сцепление, но их сегрегация происходит независимо от генов других хромосом.
Так, гетерозигота 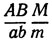 в мейозе давала бы следующие генотипы: АВМ, АВm, abM, аbm (нерекомбинанты по А и В), AbM, Abm, аВМ, аВm (рекомбинанты по А и В, образовавшиеся в результате кроссинговера). В результате потери хромосом образуются лишь генотипы
в мейозе давала бы следующие генотипы: АВМ, АВm, abM, аbm (нерекомбинанты по А и В), AbM, Abm, аВМ, аВm (рекомбинанты по А и В, образовавшиеся в результате кроссинговера). В результате потери хромосом образуются лишь генотипы
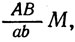
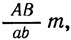
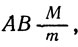
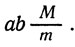
Отсутствие рекомбинации генов А и В указывает, что оба они расположены в одной хромосоме.
У некоторых плесневых грибов, например у Aspergillus nidulans, потеря отдельных хромосом нарушает течение последующих митозов. Нерасхождение хромосом и их потеря повторяются, в результате чего последовательно теряются хромосомы одного из наборов до тех пор, пока не установится сбалансированное гаплоидное состояние. Хотя такая гаплоидизация, как и мейоз, приводит к переходу диплоидного состояния в гаплоидное, она не сопровождается рекомбинацией генов одной хромосомы и поэтому дает эффективный способ выявления сцепления генов. Генотипы, которые в приведенном выше примере будут образовываться в результате гаплоидизации, следующие:
Гаплоидизация, таким образом, может быть использована для выявления сцепления.
Пример. У Aspergillus nidulans был сконструирован диплоид, гетерозиготный по генам окраски конидий w+/w и у+/у, который образовывал зеленые споры, и по четырем мутациям ауксотрофности (ad, bi, phen, pu) и мутации медленного роста (sm). Гаплоиды были отобраны на основании изменения цвета конидий - белые (w у+ и w у) и желтые (w+у). Среди них были учтены генотипы по остальным маркерам. Найдено:
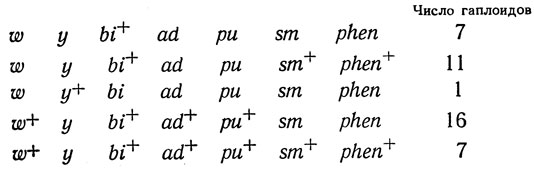
Таким образом, гены распределились по трем группам, свободно рекомбинирующимся друг с другом, но внутри которых рекомбинация не происходит:
Из этого можно заключить, что исходный диплоид был гетерозиготен по генам трех хромосом, т. е. имел генотип
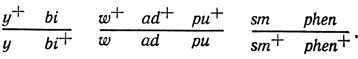
У плесневых грибов (Aspergillus, Penicillium) спонтанная гаплоидизация происходит редко. Ее частота, однако, может быть значительно увеличена при добавлении к среде культивирования n-фторфенилаланина или фунгицида бенлейта (беномила). С помощью М-фторфенилаланина удается индуцировать гаплоидизацию (с частотой 50-95 %) у Schizosaccharomyces pombe [25]. У дрожжей-сахаромицетов полная гаплоидизация при вегетативных делениях, по-видимому, не происходит, однако потеря хромосом наблюдается, что может быть использовано в генетическом анализе. Потеря хромосом эффективно индуцируется метилбензимидазол-2-ил-карбаматом (производное беномила) с частотой 2,5-6,9 % по отношению к выжившим клеткам. Для определения группы сцепления, к которой принадлежит картируемая мутация, обработке подвергают диплоид, несущий в гетерозиготе эту мутацию и соответствующие рецессивные маркеры [32].
Для повышения частоты потери хромосом используют также штаммы специального генотипа [32]. Рецессивная мутация chl1 в гомозиготном состоянии вызывает частую потерю ряда хромосом (I, III, V, VII, VIII, IX, XII, XV, XVI). Это может быть использовано для определения локализации гена либо в этих хромосомах, либо вне их, т. е. в какой-то из остальных хромосом набора.
Мутации температурочувствительности cdc6 и cdcl4 в гомозиготном состоянии приводят к тому, что гомозиготные клетки после температурного шока (36 °С, 6 ч) с высокой частотой теряют хромосомы, по одной или по нескольку сразу, что, как и в других описанных выше случаях, может быть использовано для определения локализации генов.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'