1.3.2. Митотический кроссинговер
Если потеря хромосом и гаплоидизация позволяют отнести ген к одной из групп сцепления, то изучение митотического кроссинговера дает возможность определить локализацию гена в хромосоме. Цитологическая основа митотического кроссинговера - редкая конъюгация гомологичных хромосом в митозе, сопровождающаяся обменом идентичными участками. Схема митотического кроссинговера представлена на рис. 1.4.
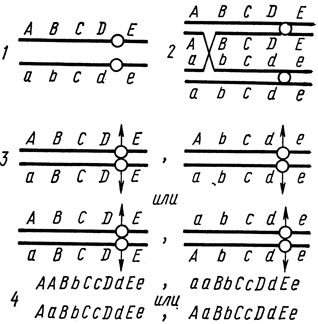
Рис. 1.4. Схема митотического кроссинговера. 1- хромосомы исходной клетки; 2 - профаза; 3 - анафаза; 4 - генотипы дочерних клеток; кружок - местоположение центромера
Если хромосома несет несколько генов в порядке ABCDOE (О - центромер), то при обмене в интервале А-В в гомозиготу переходит ген А/а с сохранением гетерозиготности в локусах В/b, С/с, D/d, Е/е. Обмен в интервале В-С переведет в гомозиготу одновременно гены А/а и В/b, а в интервале D-О - все гены этого плеча группы сцепления. Независимо от того, где произошел обмен, гены другого плеча (в нашем случае Е/е) остаются в гетерозиготе. Таким образом, А/а - самый отдаленный от центромера ген - имеет наибольшую вероятность гомозиготизации. Эта вероятность меньше для В/b и особенно для D/d. Частота выхода гена в гомозиготу характеризует его расстояние от центромера. На этом и основывается принцип картирования при митотическом кроссинговере.
Всю длину хромосомного плеча, от центромера до наиболее удаленного от него маркера, принимают равной 100 условным единицам картирования. Процент рекомбинантов по А/а, сохранивших гетерозиготность по В/b, является мерой расстояния А-В, процент рекомбинантов по A/а, гетерозиготных по С/с,- мерой расстояния А-С и т. д.
Если принять расстояние от гена А/а до центромера DОА = 100, то расстояние от гена В/b до центромера
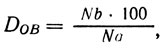
где Na - общее число гомозигот по а; Nb - число гомозигот по b среди гомозигот по а.
При митотическом кроссинговере обычно не происходят множественные обмены, которые, таким образом, не препятствуют анализу результатов. Практически задача картирования сводится к отбору рекомбинантов по самому отдаленному от центромера гену и определению их генотипов по другим сцепленным маркерам. В качестве дистального маркера удобно использовать легко выявляемый признак - селектор, позволяющий отбирать редкие рекомбинанты. Например, при работе с Aspergillus nidulans можно использовать диплоиды у+/у (образует зеленые конидии, а при гомозиготизации дает генотип у/у, проявляющийся в виде участков мицелия с желтыми конидиями) и ad/ad, su+/su (не растет на среде без аденина, а выщепляющиеся гомозиготы по рецессивному супрессору потребности в аденине генотипа ad/ad, su/su не нуждаются в этом основании и растут на минимальной среде). Отобранные по селекторам рекомбинанты испытывают по другим признакам.
Пример [16]. У аспергилла получен диплоид, гетерозиготный по маркерам su, riboA, аnА, proA, pabaA, у. Изучены генотипы диплоидных сегрегантов, образовавшихся в результате митотического кроссинговера и отобранных по признаку желтой окраски или по результату посева на среду без аденина (табл. 1.10).

Таблица 1.10. Число сегрегантов разного генотипа при митотическом расщеплении у A. nidulans
Результаты показывают, что гены su, riboA, аnА находятся в одном плече хромосомы, а у, pabaA, pro А - в другом, отсутствие сегрегантов генотипа su/su an А/an А и у/у pro А/ pro А указывает на порядок su - riboA - аnА - О и у - pabaA - рrоА - О. Принимая расстояния su - О и у - О за 100, рассчитываем расстояние до центромера остальных генов:
riboA: 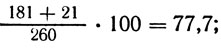
аnА: 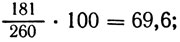
pabaA: 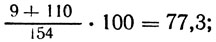
рrоА: 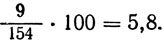
Отсюда получаем карты
su - 22,3 - riboA - 8,1 - anА - 69,6 - О; у - 22,7 - pabaA - 71,5 - proА - 5,8 - О.
Спонтанная частота митотического кроссинговера - 10-3-10-5 на деление. Она может быть увеличена на несколько порядков при облучении и при действии мутагенов.
У немногих грибов частота митотической рекомбинации оказывается необычайно высокой. Это обнаружено для несовершенного гриба фитопатогена Verticillium albo-atrum. Биологические особенности этого гриба позволили разработать особый метод митотического анализа, так называемый фиалидный анализ [21, 22]. Схема его представлена на рис. 1.5. На нем показаны конидиофор Verticillium и (при большом увеличении) отдельная веточка конидиофора, или фиалида. При делении ядра фиалиды одно из дочерних ядер остается в ней, другое переходит в отпочковывающуюся конидию; так происходит несколько раз. При выделении отдельных конидий одной фиалиды (нижняя часть рис. 1.5) можно получить клоны, нестабильные по какому-либо морфологическому маркеру (очевидно, гетерозиготные диплоиды) и стабильные (гомозиготные, проявляющие как доминантный, так и рецессивный признак). Они являются результатом реципрокной рекомбинации - митотического кроссинговера. Число конидий разного генотипа в одной фиалиде дает возможность определить момент, когда произошел кроссинговер. Фиалидный анализ оказался полезным при картировании генов.
![Рис. 1.5. Схема фиалидного анализа у Verticillium albo-atrum [16]: а - конидиофор и отдельная фиалида; б - схема последовательных делений ядер и колоний, образующихся из отдельных конидий; 1 - ядро фиалиды; 2 - ядра конидий; 3 - колонии, образовавшиеся из конидий; X - момент кроссинговера](pic/000039.jpg)
Рис. 1.5. Схема фиалидного анализа у Verticillium albo-atrum [16]: а - конидиофор и отдельная фиалида; б - схема последовательных делений ядер и колоний, образующихся из отдельных конидий; 1 - ядро фиалиды; 2 - ядра конидий; 3 - колонии, образовавшиеся из конидий; X - момент кроссинговера
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'