6. Совсем немного статистики
Теперь необходимо ввести понятия, очень важные в генетике: фенотип и генотип. Под фенотипом мы понимаем внешний вид организма, причем термин "внешний вид" употребляется в самом широком смысле. Так, фенотип бактерии - это не только ее форма и размер, но также способность бактерии вызывать заболевание, устойчивость или чувствительность к лекарствам, скорость размножения, необходимые ей питательные вещества и т. д. Фенотип человека включает тысячи физических и психических признаков: рост, частота пульса, мировоззрение, умственное развитие (если говорить только о некоторых из них, взятых наугад). Под генотипом мы понимаем весь набор генов данного организма, включая и расположение генов в хромосомах. Исчерпывающее описание фенотипа организма может потребовать практически слишком много времени и усилий, но теоретически это осуществить можно.
Подробно описать генотип даже простейшего организма невозможно потому, что каждый генотип содержит множество генов, само существование которых нам не известно. Если бы ген не был представлен различными аллеломорфами, мы не знали и не могли бы знать, что он существует. Мы знаем, например, что у крупного рогатого скота есть гены, от которых зависит, вырастут ли у данной особи рога или нет; при скрещивании рогатой и комолой пород оказывается, что эти противоположные признаки расщепляются в потомстве. Можно предположить также, что крупный рогатый скот обладает генами, которые определяют, будет ли у потомства сердце; но поскольку мы не можем скрестить двух животных, у одного из которых отсутствовало бы сердце, существование таких генов остается всего лишь догадкой. Наличие у хламидомонады генов, управляющих образованием жгутиков, смогли обнаружить только после того, как были найдены особи без жгутиков для опытов по скрещиванию, Даже у таких генетически наиболее изученных организмов, как дрозофила или кукуруза, известна лишь небольшая часть всех генов.
На практике мы никогда не имеем дела с фенотипом или генотипом как с чем-то целым. В простейших случаях ограничиваемся лишь фенотипическим эффектом отдельных генов, например влиянием пары аллеломорфных генов, которые у хламидомонады определяют образование жгутиков, или пары аллеломорфов, определяющих у гороха образование гладких или морщинистых семян. В этих случаях фенотип проявляется сразу. Но в то время как у гаплоидной хламидомонады генотип можно определить непосредственно по фенотипу, у диплоидного гороха это не так просто. У диплоида мы сталкиваемся с понятием, которое Мендель называет доминантностью против рецессивности и которое сейчас следует рассмотреть с точки зрения действия генов.
Как мы уже выяснили, диплоидный организм может быть гетерозиготным или гомозиготным по отношению к любой паре генов (см. гл. 5). Растение гороха может быть гомозиготным по "гладким" генам, гомозиготным по "морщинистым" семенам или гетерозиготным, несущим "гладкий" ген в одной хромосоме и "морщинистый" ген в соответствующем положении хромосомы-партнера (термины взяты в кавычки, так как они характеризуют форму семян, а не генов). На рис. 12, однако, мы изобразили гены так, как будто они сами по себе выглядят как признаки, вызываемые ими на семенах. На рис. 12 показаны фенотипы семян, соответствующие трем возможным генотипам формы семян. Ясно, что гомозиготы по морщинистому гену будут иметь морщинистые семена, а гомозиготы по гладкому гену - гладкие семена. Однако не очевидно, каков будет фенотип гетерозиготы. Он определяется относительной активностью влияния двух аллелей на развитие организма, и это можно выяснить путем наблюдения. В данном случае гладкий ген настолько более сильный партнер, что гетеро-зигота фенотипически неотличима от гомозиготы с гладким геном. Используя терминологию Менделя, мы назовем гладкий ген доминантным, а морщинистый ген - рецессивным. Оба термина связаны друг с другом и относятся к паре аллелей, в применении к единичному гену они не имеют смысла.
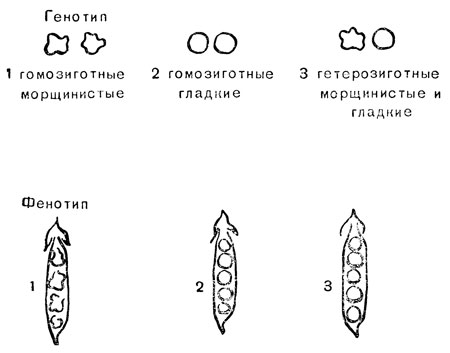
Рис. 12. Доминирование гладких над морщинистыми семенами у гороха
Подобно тому, как Джон может быть сильнее Билла, но слабее Джима, так и ген может быть доминантным по отношению к одному аллелю, но рецессивным по отношению к другому. В качестве общего вывода можно сказать, что ген А является доминантным по отношению к своему рецессивному аллелю а, когда фенотип гетерозиготы Аа определяется исключительно геном А. По этому определению условно обозначают аллели одной и той же буквой, причем доминантный член пары обозначают прописной буквой, а рецессивный член - строчной.
Из рис. 12 видно, что доминантность - это фактор, который может маскировать генотип в фенотипе; так, фенотипически семена растений 2 и 3 неразличимы, хотя генотипически они различны. Далее будет показано, как обойти эту трудность в тех случаях, когда важно установить генотип организма. Но сначала надо закончить наш экскурс в цитологию и современную генетику объяснением (используя представление о доминантности и рецессивности генов) того, как Мендель получил соотношение 3:1.
Следует вспомнить (обратимся опять к рис. 1), что скрещивание растений гороха с гладкими семенами и растений с морщинистыми семенами дает гибридные только гладкие семена, но растения, выросшие из этих семян, дают и гладкие и морщинистые семена в отношении 3 гладких к 1 морщинистому. На рис. 13 этот эксперимент воспроизведен еще раз, но с добавлением генотипов. R обозначает ген гладких семян, r - ген морщинистых. Родительские растения диплоидны и гомозиготны либо по гену R(RR), либо по гену r(rr). Их гаметы гаплоидны и несут один из двух генов: R в разновидности с гладкими семенами, а r в разновидности с морщинистыми семенами.
На рис. 13 изображен случай, когда скрещивание произошло при оплодотворении растения с морщинистыми семенами пыльцой от растения с гладкими семенами; если бы скрещивание проводили в обратном направлении, результат был таким же. Оплодотворенная семяпочка - диплоид, но в противоположность обоим родителям она гетерозиготна (Rr), так как кроме собственного гена r получила ген R через пыльцу. Семяпочка развивается в гетерозиготное (или гибридное) семя, которое вследствие доминантности R над r имеет гладкую форму семян (так же у отцовского растения).
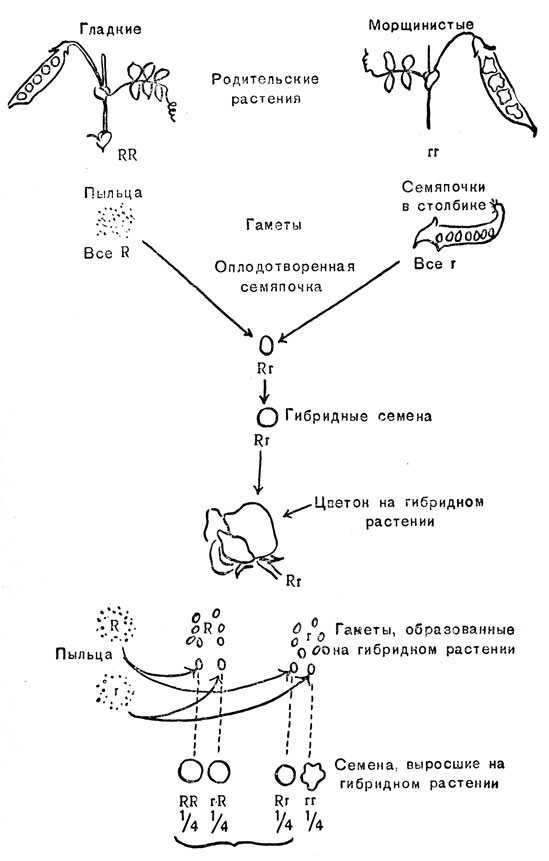
Рис. 13. Опыты Менделя, доказывающие соотношение 3:1
Когда из семян развивается растение, оно оказывается гибридом с цветами, которые, конечно, также гетерозиготны. В этих цветах при подготовке к образованию гамет протекает мейоз. Оба аллеломорфа расщепляются, и каждая гамета - пыльца или семяпочка - будет содержать или ген R, или R, но никогда оба. В отличие от того, что происходит у микроорганизмов, четыре клетки, образовавшиеся при мейозе, не остаются вместе, но судьба их в женских и мужских организмах различна. Пыльцевые зерна основательно перемешиваются, и за оплодотворение данной семяпочки соревнуется большое число мужских клеток. Семяпочки развиваются только из одной клетки из четырех, а другие три дегенерируют. Такова обычная судьба женских гамет у высших организмов, это происходит вследствие необходимости обеспечивать достаточным питанием эмбрион, который будет развиваться из оплодотворенного яйца. Возникает вопрос: что же стало в этих условиях с простым соотношением 2:2 или 1:1, наблюдаемым при расщеплении? Ответ таков: оно сохранилось, но скорее как статистическое, а не абсолютное соотношение. Легко понять, почему это так. Рассмотрим сначала мужские зародышевые клетки, а именно пыльцевые зерна у растений, соответствующие сперматозоидам животных.
Первоначально они собраны в пучки по четыре клетки, две из которых принадлежат к одному типу, две - к другому. Если взять для подсчета большое число таких пучков, общее соотношение между двумя типами все же составит 1:1 или будет весьма близким к нему. Теперь рассмотрим, женские половые клетки, т. е. семяпочки у растений и яйцеклетки у животных. Каждая из них представляет собой клетку, выжившую из четырех, которые состояли из двух клеток одного типа и двух - другого. Какая именно из четырех выживет - дело случая, причем вероятность выживания одинакова для клеток обоих типов.
Большое число женских гамет будет поэтому содержать оба типа приблизительно в одинаковом соотношении. Таким образом, примерно около половины пыльцевых зерен и половины семяпочек, образованных гибридным растением Rr, будет нести R ген, а другая половина - R ген. Оплодотворение может произойти четырьмя различными путями с равной вероятностью: пыльца R может оплодотворить R или R семяпочки, а пыльца R - R или R семяпочки. В результате потомство (семена) будет состоять из трех различных генотипов: гомозигот Rr и Rr и гетерозигот Rr. Гетерозигота может образоваться двумя различными путями в зависимости от того, который из двух аллелей будет доставлен пыльцой и который - семяпочкой; поэтому гетерозигота образуется вдвое чаще, чем гомозигота. Окончательное соотношение трех генотипов: 1/4Rr : l/2Rr : 1/4Rr или 1Rr:2Rr:1Rr. Однако это соотношение проявляется не столь явно из-за доминантности гена R. Фенотипически Rr и Rr семена неразличимы, и отношение между фенотипами равно 3:1.
Если взглянуть на цифровые фактические данные, действительно полученные Менделем, то окажется, что они весьма близки к соотношению 3:1, но не абсолютно совпадают. Этого следовало ожидать, исходя из статистической природы этих данных.
Теперь понятно, почему я назвала соотношение 3:1 скорее отдаленным последствием решающего соотношения в результате расщепления гамет, так как для того, чтобы оно осуществилось, требуется наличие особых условий: 1) один аллель должен полностью доминировать над другим; 2) скрещивание должно быть или в результате самооплодотворения гибрида, или оба родителя должны быть гетерозиготами по одной и той же паре аллелей. Следующие два примера показывают, что происходит, если какое-либо из этих условий не выполняется. Полная доминантность у гетерозигот далеко не всегда является правилом. Часто доминантность выражена неполно, и фенотип гетерозиготы представляет собой нечто среднее между двумя гомозиготами. Чаще всего это наблюдается для так называемых количественных признаков, вроде роста или веса, однако и качественные признаки (например, наличие или отсутствие окраски) также могут проявлять неполную доминантность. У породы крупного рогатого скота - шортгорнов интенсивность окраски определяет пара аллелей. Одна гомозиготная особь полностью окрашена, другая - белая. Гетерозиготная особь - смесь белых и пигментированных волос. Если основная окраска шкуры, управляемая другой парой генов, рыжая, то такая смесь дает чалую окраску. На рис. 14 показан результат скрещивания чалого быка и чалой коровы. Оба аллеля, так же как и на рис. 13, обозначены буквами R и R, так что результаты обоих скрещиваний легко сопоставить друг с другом. Скрещивание двух чалых животных соответствует самооплодотворению у гибридного растения гороха, расщепление на два типа сперматозоидов и яйцеклеток соответствует расщеплению двух типов пыльцы и семяпочек; потомство состоит из трех генотипов: Rr, Rr и Rr в отношении 1:2:1. При отсутствии доминантности одного аллеля над другим соотношение фенотипов такое же, как и генотипов, а именно 1/4 рыжих, 1/2 чалых и 1/4 белых телят. Нельзя, естественно, ожидать, что такое соотношение обнаружится у первых же четырех или восьми телят, родившихся от такого скрещивания; однако, если объединить большое число данных по нескольким стадам, получается хорошее совпадение фактических данных с теоретическими.
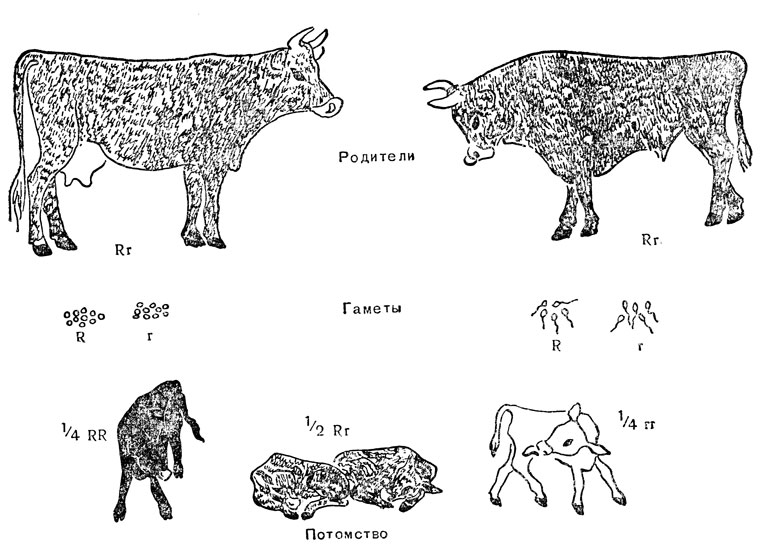
Рис. 14. Наследуемость чалой окраски у шортгорнов; соотношение 1:2:1
На основании построенной им гипотезы Мендель предсказал, что все гладкие семена в первом поколении и 2/3 их во втором поколении будут гибридными, или, как мы можем теперь их назвать, гетерозиготными. Для проверки Мендель выращивал растения обоих поколений и использовал пыльцу для опыления растений, выращенных из морщинистых семян. Скрещивания, полученные с гибридами первого поколения, и 2/3 скрещиваний, полученных с гибридами второго поколения, дали ожидаемые результаты (рис. 15). Гибридные растения дали два типа пыльцевых зерен, R и R, в равных количествах, тогда как растения, выращенные из морщинистых семян, дали только R семяпочек. В результате оплодотворения образовалось одинаковое количество гладких и морщинистых семян. Такой вид скрещивания, при котором гибрид скрещивается с одной из родительских линий, называется обратным скрещиванием. Тогда, как в данном примере, родительский штамм рецессивного типа, соотношение фенотипов в потомстве точно такое же, как соотношение гамет при расщеплении, а именно 1:1, Это соотношение и результат обратного скрещивания имеют большое значение, чем представляющее лишь исторический интерес отношение 3:1, наблюдаемое при скрещивании двух гетерозигот. Применение обратного скрещивания для выведения новых пород животных будет обсуждаться в следующей главе.
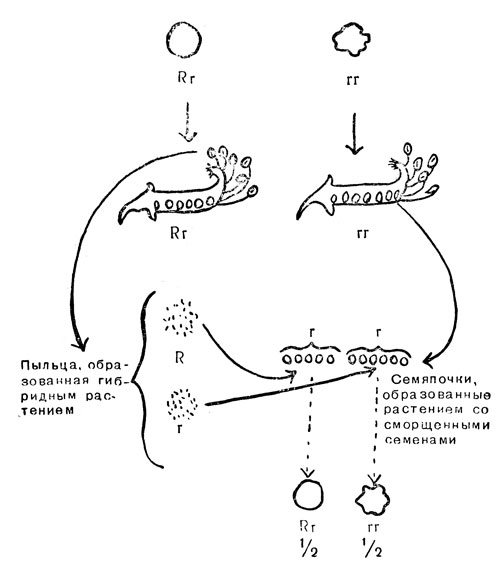
Рис. 15. Обратное скрещивание гетерозиготного гороха с гомозиготным рецессивным
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'