7. Об овцах, норках и собаках
В 1791 г. на одной ферме штата Массачусетс в США одна овца принесла ягненка с длинным туловищем и короткими искривленными ногами. Хотя отнятый у матери ягненок был менее приспособлен к жизни, чем другие, впоследствии он вырос в здорового барана и от него пошло похожее на него потомство. Так было положено начало выдровой, или анконской, породе овец. Внешне эти животные производили столь малоприятное впечатление, что Ч. Дарвин назвал их "полууродами", а один специалист по овцеводству в Нью-Йорке писал в 1809 г.: "Если цивилизованная нация, которая обладает представлением о стандарте красоты, дает согласие на уродование божьих созданий и воздвигает уродству алтарь, принося в жертву радости жизни беспомощных и полезных животных, стоит ли удивляться, что дикари, не знающие красоты пропорций и гармонии форм, пытались обуздать причиняющую хлопоты подвижность своих овец, поощряя разведение овец с уродливыми хвостами, подобно тому, как некоторые из нас предпочитают у них рахитичные ноги"?
В этом вопросе уже содержится намек на причины, объясняющие, почему фермерам нравились анконские овцы, несмотря на их уродство: из-за своих коротких ног эти овцы не могли перепрыгивать через низкие изгороди пастбищ и поэтому не могли наносить ущерб полям; отпадала необходимость в пастухах и собаках. Анконская порода была с легкостью выведена и легко сохранялась.
При скрещивании барана - родоначальника анконской породы с его матерью было получено потомство также анконской породы, и при их дальнейшем скрещивании получались только анконские овцы, впоследствии все скрещивания двух анконов давали ягнят только анконской породы.
С другой стороны, у нормальных родителей могут рождаться анконские ягнята, и раз это произошло однажды, может произойти и снова, хотя большая часть ягнят от подобных скрещиваний бывает нормальной.
Ч. Дарвин в книге "Изменчивость животных и растений в одомашненном состоянии" упоминает об анконских овцах, как о редком случае внезапного возникновения новой породы. Хотя основной труд Г. Менделя появился несколькими годами раньше выхода этой книги, Дарвин не был знаком с работами Менделя. Зная их, он, возможно, пришел бы к простому объяснению возникновения анконской породы.
Породы обычно отличаются друг от друга многими генами, и поэтому для их выведения необходимо проводить отбор на протяжении многих поколений, анконские же овцы отличаются от первоначальной породы только одним геном, вызывающим своеобразное строение их тела. Как показано на рис. 16, I, этот ген может быть передан потомству от внешне нормальных родителей, которые, следовательно, должны быть гетерозиготными по данному гену. Согласно принятому определению, ген, скрытый в гетерозиготном состоянии, носит название рецессивного. Будучи рецессивным, ген анконской породы проявляется только в гомозиготах; таким образом, все анконские овцы являются гомозиготными и приносят только анконских ягнят (рис. 16, II).
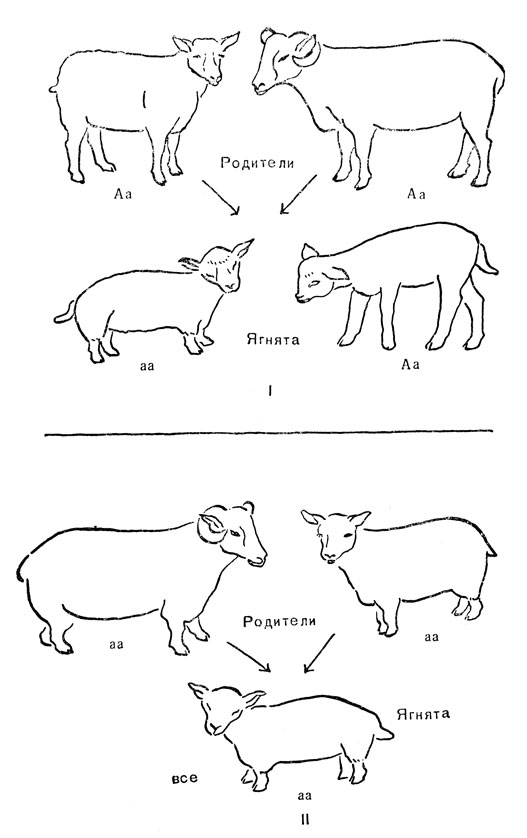
Рис. 16. Наследуемость анконского признака у овец: I - у нормальных родителей родился один анконский ягненок и один нормальный ягненок; II - родители-анконы рождают только ягнят-анконов; а - ген анконского признака; А - его нормальный аллеломорф
В менделевском объяснении происхождения анконской породы остается без ответа вопрос о возникновении анконcкого гена. Почему анконские овцы не появились до 1791 г. даже на ферме, где два гетерозиготных родителя дали первого анконского ягненка? Вопрос этот сложный. Можно ответить лишь формально, дав название редкому и все еще не до конца понятному процессу, при котором давно создавшийся ген образует новый аллеломорф. Этот процесс называется мутацией. Подробнее мы будем говорить о нем ниже. Сейчас достаточно сказать, что анконский ген возник в определенном стаде и в определенное время в результате мутации одного из генов, который у овцы управляет развитием скелета. Но по существу мы не можем быть уверены относительно точного времени возникновения гена. Поскольку он рецессивен, то вполне мог переходить в скрытом виде из поколения в поколение, прежде чем спаривание двух гетерозигот привело к появлению первого анконского ягненка. Мутировавший ген подобно своему предшественнику воспроизводит новые гены по своему подобию, и потомки первого мутировавшего гена имелись у всех анконских овец, разводимых в Новой Англии (США) до второй половины девятнадцатого столетия.
В начале нашего столетия анконская овца появилась в Норвегии. Сказать трудно, был ли этот европейский анконский ген привезен в Норвегию в гетерозиготном состоянии или же возник в результате новой мутации.
В то время как в практике сельского хозяйства использование одной единственной мутации для выведения новой породы встречается редко, это очень распространено среди любителей разводить животных для забавы. В отличие от селекционеров, интересующихся плодовитостью, крепостью сложения, молочной продуктивностью или яйценосностью или другими свойствами, зависящими от действия многих генов, любители интересуются внешним видом животных, который может довольно резко изменяться в результате мутации одного-единственного гена. Примером этому служит возникновение новых пород норки. Разведение норки с коммерческими целями началось сравнительно недавно. Первые большие выставки норок были устроены в 1929 г. в США и в 931 г. в Канаде. Животные на этих выставках относились по окраске к типу темной окраски, характеризующей диких норок. В 1931 г. хозяин одной висконсинской фермы обнаружил у себя самку норки светлого тона.
После спаривания этой самки с самцом темной окраски все потомство получилось темным, но при обратном скрещивании одного из самцов со своей матерью получили более светлоокрашенных норок. При спаривании друг с другом животных этого типа, называемых с тех пор серебристо-голубыми, или платиновыми, норками, все потомство оказывалось похожим на своих родителей, и таким образом получилась гомозиготная порода платиновой норки.
Читателю должно быть ясно, что история возникновения атиновой норки - почти точное повторение истории ансонской овцы, причем и в том и в другом случае возникновение новых свойств обусловлено рецессивным геном, появившимся в результате мутации. Между прочим, сходная мутация возникла через несколько лет на другой норковой ферме, и ее использовали для получения чистокровного отродья. При скрещивании животных обоих отродьев в потомстве получали норок платиновой окраски. Для генетиков этот факт служит доказательством того, что обе мутации были идентичными. Эти доказательства, возможно, читателю не понятны; к этому вопросу мы вернемся в гл. 22.
Заводчик, желающий ввести платиновую мутацию в своем стаде, может сделать это путем случайных скрещиваний, однако есть возможность достичь того лее результата с меньшей затратой времени и средств, если применить на практике принципы Менделя (рис. 17).
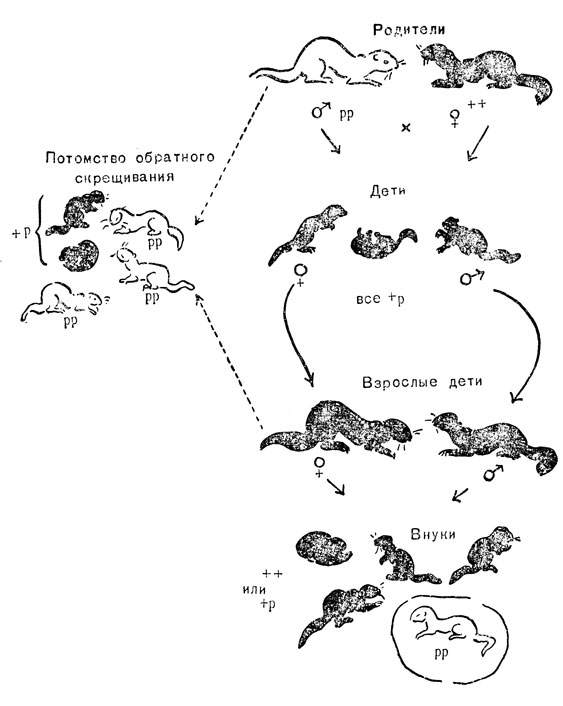
Рис. 17. Наследуемость платиновости меха у норок: р - ген платиновости; + - его нормальный аллеломорф
Рецессивный платиновый ген обозначен буквой р, его темный аллель, имеющийся у диких норок, знаком "+". Часто бывает удобным обозначать знаком "+" гены, имеющиеся у диких животных, это относится как к доминантным, так и рецессивным генам. Очевидно, что, если как в разбираемом примере, мутантный ген является рецессивным партнером, аллель, имеющийся у дикой разновидности, должен быть доминантным.
Таким образом, платиновых животных генетически обозначают рр; особи дикой разновидности - "+ +"; гетерозиготы - +р (фенотипично дикой окраской). При скрещивании платинового самца (рр) с дикой самкой (+ +) все сперматозоиды несут ген р, все яйцеклетки - ген "+", и все потомство имеет генотип +р и темный фенотип. При скрещивании их друг с другом половина яйцеклеток и половина сперматозоидов несет ген р, поэтому примерно одна четверть потомства получит этот ген и со сперматозоидом, и с яйцеклеткой и будет характеризоваться платиновой окраской. Более совершенным приемом получения норок платиновой окраски надо считать скрещивание платинового самца с его гетерозиготными (+р) дочерьми, так как при этом все сперматозоиды и половина яйцеклеток будут нести в себе желаемый ген, и половина потомства получится платинового типа. Лишь только будут получены платиновые животные обоих полов, их можно использовать для образования и чистопородной платиновой линии.
Платиновая норка была первой мутантной норкой. Она произвела сенсацию на аукционе в Нью-Йорке в 1944 г., где цена за одну шкурку достигла наивысшей - 265 долларов. Вскоре появились другие мутации. Одной из наиболее знаменитых и наиболее ценимых была мутация светлой окраски с темным крестом на спине, так называемая кохинур (рис. 18). Первое животное с таким крестом передало этот признак части своего потомства, поэтому для генетиков было ясно, что в отличие от платиновой мутации мутация черного креста должна считаться доминантной по отношению к своему нормальному аллелю. При скрещивании между собой животных с черным крестом в потомстве были животные трех типов: темные, почти белые и животные с черным крестом, причем последних было почти вдвое больше, чем животных каждого из первых двух типов. Ситуация в точности такая же, как у скота чалой породы (см. рис. 14). Если обозначить мутантный ген S а нормальный аллель знаком "+", то гетерозигота S+ будет нести черный крест, а две гомозиготы "+ +" и SS будут соответственно темной и светлой окраски. Практическим результатом неполного доминирования S над "+" является то, что кохинуровские норки, так же как и чалые коровы и быки, никогда не могут быть получены в виде гомозиготной, чисто размножаемой породы. Селекционеры вынуждены выбирать между приплодом, являющимся смесью кохинуровских и белых норок (см. рис. 18, I), кохинуров и темных (см. рис. 18, II) или смесью всех трех типов (см. рис. 18, III).
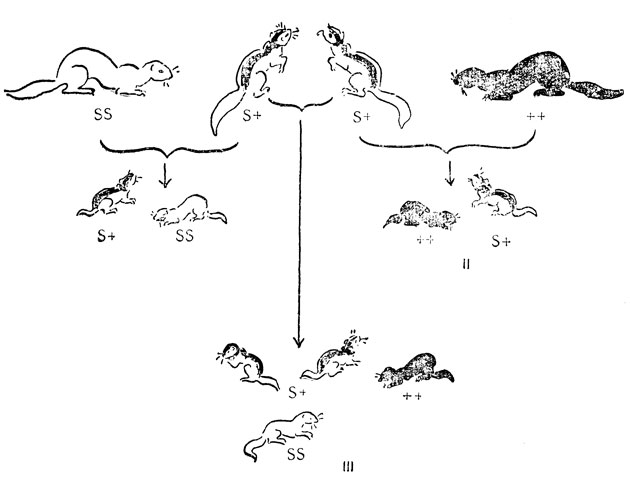
Рис. 18. Наследуемость кохинуровой окраски меха у норки: I - кохинуровая норка, скрещенная с белой; II - кохинуровая, скрещенная с темной; III - кохинуровые скрещенные между собой; S - ген белой окраски меха; + - ген темной окраски меха
С другой стороны, неполная доминантность имеет то преимущество, что гомозиготных особей доминантного типа можно с легкостью отличить от гетерозиготных; например, очень легко вывести отродье чистокровных белых норок путем скрещивания белых (SS) между собой. При полной доминантности это обстоит не так; в этом случае, чтобы отличить гетерозиготу от доминантной гомозиготы, необходимо провести испытания по потомству.
У некоторых пород собак черная окраска шерсти В доминирует над рыжей b, и если рыжая мутация вкраплена в черную породу, она будет распространяться по питомнику, так что среди потомства двух черных родителей Bb могут неожиданно появиться рыжие щенки bb.
Владелец собачьего питомника, желающий избавиться от гена рыжей окраски, может сделать это наиболее эффективно, используя рыжих животных в качестве пробных присутствие рыжего гена у самок и самцов с черным фенотипом (рис. 19). Когда гетерозиготная особь Bb спаривается с рыжей особью bb, в потомстве рано или поздно должен появиться рыжий щенок. Гомозиготная особь черного типа BB, скрещенная с рыжей, даст потомство, состоящее из одних только черных щенков Bb. Поэтому животные с черной окраской, давшие при таких скрещиваниях шесть или семь черных щенков и ни одного рыжего, могут использоваться для восстановления чистокровной черной породы.
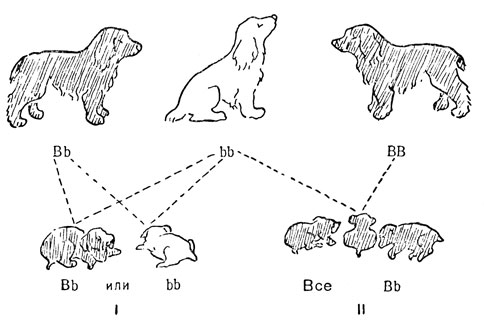
Рис. 19. Испытания по потомству на присутствие рецессивного гена, вызывающего рыжую окраску шерсти у собак: B - ген черной окраски; b - ген рыжей окраски
Эти простые испытания по потомству на присутствие нежелательных генов могут быть выполнены всегда, когда в качестве пробного животного используется рецессивная гомозиготная особь (рыжая в приведенном выше случае). К сожалению, применительно к большинству действительно вредных генов, как мы убедимся в следующей главе, дело обстоит не так.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'