23. Гены и признаки
Вряд ли можно найти хоть один признак в любом организме, который не находится в некоторой степени под влиянием генотипа. Гены управляют окраской и формой растений и животных, способностью животных видеть, слышать и обонять, определяют сопротивляемость человеческого организма к патогенным бактериям и способность бактерий заражать человека. Они влияют на темп роста теленка на данной диете, на способности ребенка к обучению и на вероятность нарушения психики человека под влиянием психического напряжения. Они определяют, будет ли водоросль хламидомонада иметь жгутики для плавания и будет ли дрожжевая культура расти на галактозе (см. гл. 5). Гены влияют на самые важные и самые основные жизненные процессы; они проявляют свое действие на всех стадиях развития. Действительно, само выживание от момента зачатия до глубокой старости в некоторой степени находится под влиянием генотипа. Обнаружено, что у цыплят, крыс и мышей мутантные гены убивают зародыш грубым вмешательством в развитие таких жизненно важных органов, как скелет, мозг, почки. Несомненно, что нормальные аллели этих генов влияют на нормальное развитие этих органов. Также несомненно и то, что подобные же гены влияют на развитие человеческого эмбриона.
Прослеживая жизнь организмов, мы обнаруживаем, что инбридные линии мышей отличаются средней продолжительностью жизни индивидов. Вряд ли можно сомневаться в том, что продолжительность жизни человека в какой-то степени определяется генами, хотя большая генетическая и экологическая изменчивость людей затрудняет четкое решение вопроса.
По всей видимости, большинство генов абсолютно необходимо для выживания организма. У экспериментальных животных или растений очень маленькие куски хромосом можно разрушить, например, рентгеновскими лучами (см. гл. 4, рис. 5 и 6). Попытки вырастить индивиды с нехватками одинаковых кусков у обеих хромосом-партнеров редко заканчиваются успешно; какого бы куска хромосомы ни хватало, в нем почти всегда оказываются гены, без которых выживание невозможно. На примере дрозофилы показано, что присутствие таких, казалось бы, несущественных генов, от которых зависит окраска глаз и форма краев крыльев, необходимо хотя бы в одной хромосоме для обеспечения развития зародыша до стадии окрыленной особи. Если бы эти гены оказывали лишь незначительное влияние на внешние признаки, этот факт было бы трудно понять. Однако имеются убедительные доказательства того, что большинство генов, если не все, действуют на многие признаки. Этот вывод получен в результате изучения мутантных генов.
В существовании нормального гена можно убедиться только после хотя бы одного его мутирования, а его действие оценивается только путем сравнения нормальных особей и мутантов. Многие мутантные гены одновременно оказывают влияние на разные признаки. У дрозофилы, несущей мутантный ген lozenge, глаза слипаются и имеют вид, похожий на поверхность кипящих сладких веществ, исчезают коготки и подушечки на лапках, так что мухи оказываются больше не в состоянии подниматься по стеклянной стенке банки, и самки делаются бесплодными. Японские вальсирующие мыши лишены способности слышать, кудрявые куры (см. гл. 16) имеют недоразвитые яичники и несут мало яиц. Правильно считать, что нормальные аллеломорфы этих генов необходимы для нормального развития глаз, лапок и половых органов у дрозофилы; органа слуха и нервной системы у мышей; оперения и яичников у домашней птицы. При любом тщательном изучении мутантного гена удается обнаружить, что он оказывает многостороннее действие. Большинство мутантных генов снижает жизнеспособность, поэтому не удивительно, что их нормальные аллели необходимы для жизни. Способность гена оказывать много различных эффектов называется плейотропией, или многонаправленностью. Из сказанного выше ясно, что плейотропия является правилом, а не исключением.
Подобно тому как каждый ген влияет на многие признаки, каждый признак определяется многими генами; это явление известно под названием "взаимодействия генов". Кстати, взаимодействие генов и плейотропия дополняют друг друга; вывод, что многие гены влияют на жизнеспособность плейотропически, можно выразить по-другому, сказав что жизнеспособность находится под влиянием многих генов. Аналогично этому многие мутантные гены и, следовательно, их нормальные аллели влияют на плодовитость, и наоборот, плодовитость зависит от действия всех этих генов, так же как и других, не оказывающих бросающегося в глаза действия на внешние признаки. Даже очень специфические признаки, такие, как цвет радужной оболочки глаз человека, зависят от действия многих генов.
На рис. 72 показано взаимодополняющее соотношение между плейотропией и взаимодействием генов для некоторых мутантных генов домашней мыши. Чтобы избежать мелких деталей, были взяты "признаки" в широком плане, и их следует лучше называть "органами" или "функциями". Выбор более мелких признаков, таких, как цвет глаз или строение хвоста, дал бы точно такую картину. Линия, соединяющая ген с признаком, означает, что ген влияет на этот признак. То, что от каждого гена идет несколько линий иллюстрирует явление плейотропии; то, что несколько линий оканчивается на каждом признаке, иллюстрирует взаимодействие генов. Диаграмма сознательно сделана простой, изображающей только несколько генов и признаков. Если взять больше признаков на правой стороне рисунка, большее число линий начиналось бы совместно в большинстве генов слева. Если изобразить больше генов слева, больше линий сошлось бы у многих признаков на правой стороне.
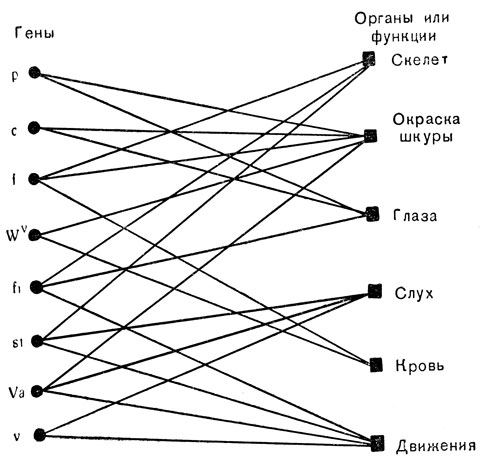
Рис. 72. Гены и признаки у домашней мыши: p - красный; с - альбинос; f - согнутый хвост; Wv - доминантная пятнистость; fi - беспокойный; st - трясучий; Va - переваливающийся; v - вальсирующий
Беспорядочный характер пересечения линий друг с другом обозначает, что система признаков, контролируемых одним геном, варьирует у разных генов. Так, все гены fi, st, Va и v вызывают нервные расстройства, проявляющиеся в виде круговых и дрожательных движений; но мышь с геном fi может слышать, в то время как три другие мутанта глухие; только ген Va вызывает неравномерную окраску шкурки, ген fi оказывается причиной образования изъязвлений на глазах и только st укорачивает хвост. Следует опять вспомнить, что эти взаимодействия между мутантными генами и признаками указывают на некоторые пути действия нормальных аллелей в процессе нормального развития.
Нам предстоит познакомиться с несколькими примерами плейотропии (т. е. способностью оказывать влияние одновременно на несколько признаков) и взаимодействия генов. После этого нам станет ясно, как груба и упрощенна была схема на рис. 72.
Плейотрэпия: один ген влияет на несколько признаков
Серповидная анемия. В 1910 г. молодой негр из Вест-Индии пришел на прием к американскому врачу по поводу лихорадки. Так как он жаловался на недомогание в течение длительного времени, врач произвел тщательное обследование и обнаружил, что молодой человек страдал резкой анемией неизвестного типа. При исследовании под микроскопом его кровь представляла поразительную картину (рис. 73, а). В то время как у здорового человека эритроциты (красные кровяные тельца) все одинаковой округлой формы, в крови этого больного было много эритроцитов серповидной формы.
Несколько лет спустя такая же ненормальность была обнаружена в крови отца и сына, страдавших анемией. Заболевание назвали серповидноклеточной анемией; оно четко отличается от обычных форм анемий. Как только это привлекло всеобщее внимание, все чаще стали обнаруживать подобные случаи, вначале исключительно среди негров. Вскоре стало ясно, что заболевание было в какой-то степени обусловлено наследственностью, но его правильную генетическую основу выявили только после появления методики, давшей возможность точно устанавливать явление серповидности. Эта методика заключается в сохранении образца крови изолированно от воздуха в течение дня или более. За это время кровяные клетки больного приобретают причудливую форму, изображенную на рис. 73, б. С помощью этой методики установили, что наряду с типичной картиной крови, обнаруживаемой у больных, страдающих серповидноклеточной анемией, нерезкие, но ясно различимые серповидные формы клеток наблюдаются в крови примерно 9% американских негров, не страдающих анемией. Эти нерезкие доброкачественные серповидные изменения особенно часто встречаются среди родственников больных, страдающих серповидноклеточной анемией, в частности их всегда обнаруживают у обоих родителей больного ребенка. Это привело к концепции, полностью подтвержденной в результате дальнейших исследований, что заболевание вызывается геном S, который в гетерозиготном состоянии S+ дает мягкие формы серповидности, не сопровождаемые анемией, в то время как гомозиготы по SS дают анемию.
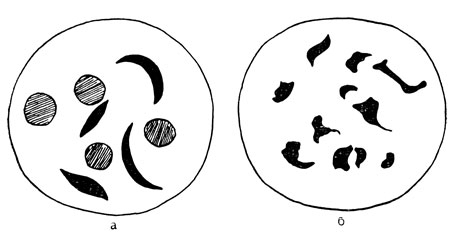
Рис. 73. Клетки крови у больного серповидноклеточной анемией (а). Серповидность особенно выявляется после пребывания крови в изолированном от воздуха состоянии (б)
Лабораторные исследования измененной крови выявили, что ген поражает гемоглобин, железосодержащий пигментный белок эритроцитов. В электрическом поле, подобном изображенному на рис. 35, молекулы гемоглобина SS-индивидов двигаются более медленно по сравнению с молекулами гемоглобина нормальных людей.
Кровь гетерозигот S+ содержит оба вида гемоглобина в приблизительно равных пропорциях; по-видимому, один вид формируется под влиянием гена S, другой - под влиянием его нормального аллеля. Это как раз является хорошей иллюстрацией относительности терминов "доминантный" и "рецессивный": ген может быть доминантным на одном уровне наблюдения и рецессивным - на другом. На уровне химического изучения гемоглобина ген S и его нормальный аллеломорф действуют независимо друг от друга, подобно генам групп крови A и B (см. гл. 10); на уровне изучения микроскопической картины крови ген S дает S неполное доминирование над своим нормальным аллелем; и, наконец, клинически ген S рецессивен по отношению к его нормальному аллелю.
Исследования встречаемости гена S во многих странах дали любопытные результаты. Сначала казалось, что наличие этого гена присуще только неграм Африки и обеих Америк. Позднее людей с серповидными формами эритроцитов обнаружили среди итальянцев, греков и других народностей вокруг Средиземного моря и среди племен индейцев Южной Америки. Распределение этих людей носило очаговый характер. Если в определенных районах Африки их обнаруживали в большом количестве, то в других областях они практически отсутствовали. Даже в пределах одной и той же территории нередко наблюдались поразительные различия в распределении людей с серповидными клетками. Например, в Уганде число таких людей составляло половину одного племени и было равно нулю в другом.
Самым загадочным в этом исследовании было то, что обладатели серповидных эритроцитов (S+) встречались особенно часто в определенных популяциях. Гомозиготы по S редко достигают взрослого состояния и не имеют детей: природа делала их полностью стерильными. Хотя, как мы знаем из главы 13, стерилизация гомозигот по рецессивному гену очень неэффективна в смысле освобождения от данного гена, она способна поддерживать частоту его проявления на низком уровне. Действительно, другие гены, приводящие к естественной стерилизации гомозигот, например ген амавротического слабоумия, встречаются редко даже среди гетерозигот. Таким образом, создалось впечатление, что в определенных популяциях бездетность и гибель гомозигот компенсировалась тем, что гетерозиготы имели некоторое естественное преимущество, дававшее им возможность по сравнению с остальным населением иметь больше детей, половина которых наследовала бы ген S.
Возможным ключом к выяснению природы этого преимущества оказалось открытие, что обладатели серповидных форм часто встречаются в малярийных местностях и редко встречаются или совсем отсутствуют в местностях, свободных от малярии. Поэтому предположили, что ген S каким-то образом предохраняет своих носителей от малярийного паразита. Исследования по проверке этого предположения подтвердили его. Из 288 детей в малярийной местности в Африке 43 ребенка оказались гетерозиготными по S и 12 из них, т. е. 28%, были носителями возбудителя малярии в крови; из оставшихся 245 детей, не имевших гена S, 113, т. е. 46% детей, были носителями паразита. В одном африканском племени 30 добровольцев заразили малярией; 15 из них были S+ , и только двое из них заболели; остальные 15 оказались "++", и из них только один человек не заболел.
Таким образом, имеется хорошее, но еще недостаточно убедительное доказательство плейотропного действия гена S. Тот же самый измененный гемоглобин, который у гомозигот приводит к анемии, делает кровь гетерозиготных людей непригодной средой для жизни малярийного паразита. Так как малярия нарушает физическое и психическое развитие, действуя на разные механизмы, конечные плейотропные влияния гена, обусловливающего восприимчивость к этому заболеванию, будут сопровождаться далеко идущими последствиями.
Если это объяснение загадочного распространения гена правильно, то мы столкнулись со случаем, когда гетерозиготный по двум аллелям (S+) организм более жизнеспособен, чем гомозиготный по SS или "++". В главе 22 это явление рассматривалось как возможная действенная причина гибридной силы.
Наследуемость карликовости у мышей. В 1929 г. несколько карликовых мышей появилось в мышином питомнике Гарвардского университета (рис. 74). Они не размножались, но когда мыши одного с ними помета скрещивались, одна четверть потомства оказывалась карликовой. Это объясняется действием рецессивного гена. Мышь, гомозиготная по этому гену, прекращала расти в возрасте конца второй недели или немногим позже, и вес ее обычно достигал 6 - 8 г, в то время как вес нормальной мыши равнялся 20 г и более. На рис. 74 показана взрослая карликовая мышь рядом с нормальной мышью одного с ней помети. Ген карликовости дает ряд плейотропных проявлений (рис. 75). На ранних стадиях развития, даже еще до остановки роста, гомозиготных особей можно отличить от нормальных мышей того же помета по тупым мордочкам и более коротким ушкам и хвостам. В то время как нормальные молодые мыши подвижны и легко возбудимы, карликовые мыши робки и вялы. Они очень чувствительны к колебаниям температуры; однако при содержании в условиях нормальной температуры они переносят голодание лучше, чем нормальные мыши. Продолжительность их жизни короче, самцы и самки стерильны. При исследовании их внутренних органов обнаруживается, что большая часть желез внутренней секреции - щитовидная, гипофиз и гонады - меньше и ненормального строения. Известно, что гипофиз - маленькая железа в основании мозга - управляет выделением всех гормонов, поэтому его часто называют "хозяином желез внутренней секреции": некоторые ученые пытались поэтому лечить наследственную карликовость с помощью гормонов гипофиза.

Рис. 74. Карликовая мышь рядом с нормальной
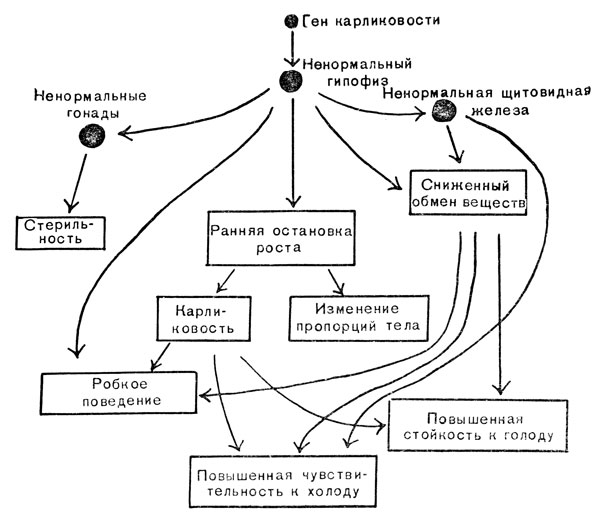
Рис. 75. Плейотропное действие карликового гена мыши
Кусочки свежей ткани гипофиза крысы подсаживали ежедневно под кожу карликовым мышам, уже остановившимся в росте. Успех лечения был изумительным. Леченые животные приобретали нормальный вес и пропорциональные размеры, становились живыми, подвижными и начинали вести себя во всех отношениях как нормальные мыши. Самцы становились способными давать потомство, самки оставались стерильными, несмотря на то что их яичники приобретали обычные размеры и строение. Все железы внутренней секреции, за исключением одной, становились нормальными. Исключением оказался гипофиз, совершенно не изменившийся под влиянием лечения. Этого можно было ожидать, если основной дефект, порождающий все остальные, затрагивает ведущую железу. Дело в том, что имплантирование гипофиза может стимулировать функцию всех подчиненных ему желез внутренней секреции, но не может устранить поражения самого гипофиза.
Взаимодействие генов: один признак зависит от многих генов
Образование хлорофилла. В клетках зеленых растений зеленые и некоторые желтые пигменты находятся внутри массы маленьких телец, называемых хлоропластами (рис. 76). Низшие растения, например водоросли, могут иметь один большой хлоропласт. До изобретения электронного микроскопа о детальном строении хлоропласта было известно мало. Теперь при помощи электронных микроскопов получают фотографии (рис. 77), выявляющие чрезвычайно сложные структуры хлоропластов. Исследования при помощи электронного микроскопа, проведенные параллельно с генетическими наблюдениями, дали много для понимания роли генов в развитии хлоропласта, и, хотя эти исследования находятся еще в зародыше, уже начала выявляться приблизительная картина взаимодействия генов.
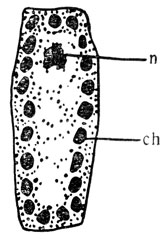
Рис. 76. Растительная клетка с ядрами и хлоропластами
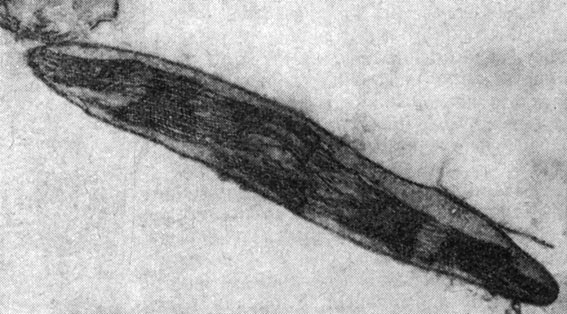
Рис. 77. Хлоропласт ячменя под электронным микроскопом
Детали этой картины еще в значительной степени неизвестны, но, по-видимому, в общих чертах картина будет похожа на ту, что изображена на рис. 78. Электронномикроскопические исследования показали, что нормальный хлоропласт V проходит стадию II - IV маленькой бесцветной гранулы I, которая в процессе развития не только изменяет размеры и форму, но также образует сложную внутреннюю сеть пластинок, между которыми взвешено некоторое количество глобул. На определенных стадиях развития хлоропласт образует пигменты, сначала желтый, потом зеленый. На диаграмме показано, что источник пигментов расположен вне хлоропласта; это сделано для большей ясности, но в действительности пигменты образуются внутри хлоропласта. Без сомнения, этими процессами управляет большое число генов, так как было найдено большое количество мутаций, которые тормозят образование хлорофилла и хлоропласта. Более ста генов выявилось в результате мутирования; вполне вероятно, что большее количество пока остается невыявленным, так как они еще не дали начала доступным для наблюдения мутациям. Хорошо также известно, что различные гены управляют различными стадиями образования хлорофилла; мутации могут приостановить развитие хлоропласта на различных стадиях и различным путем. На диаграмме показано большое количество генов (от A до G), существование которых было выявлено таким путем. Кроме того, здесь показан ген H, который не оказывает влияния на образование хлорофилла, но его присутствие оказывается необходимым для выживания уже образовавшегося хлоропласта; мутация этого гена приводит к разрушению (выцветанию) хлоропласта.
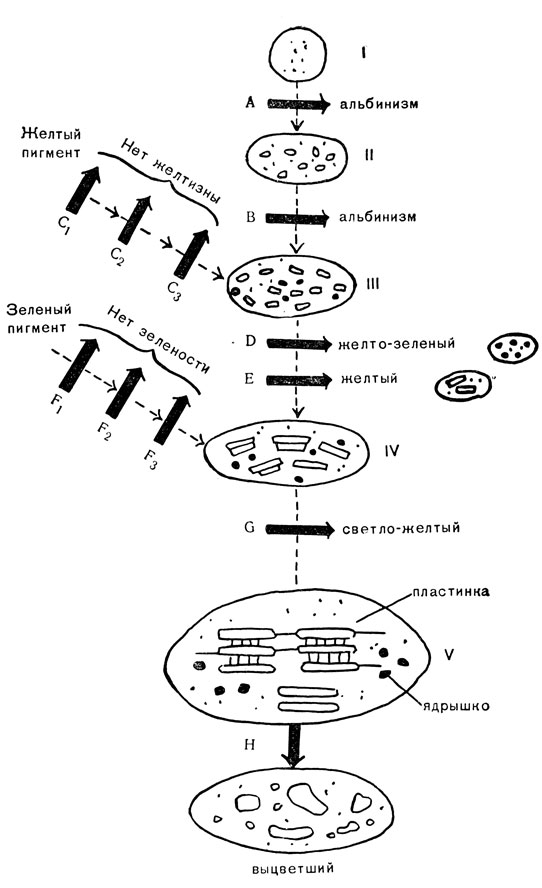
Рис. 78. Частично гипотетичная схема генетического управления синтеза хлорофилла: I - V - стадии в развитии нормального хлоропласта; A - G - гены или группы генов, ведающих развитием хлоропласта; → - блокада процессов развития; H - гены, необходимые для существования нормальных хлоропластов

Необходимо обратить внимание на следующие моменты: 1) гены, управляющие строением хлоропласта (A, B, D, E, G), действуют серийно, т. е. друг за другом; каждый ген вступает в действие лишь после того, как окончилось действие предшествующего. Так, например, ген B вступает в действие только после прекращения функции гена A; если развитие хлоропласта остановилось на первой стадии вследствие мутирования гена A, то ген B и все последующие не могут начать действовать; 2) на диаграмме показано, что то же самое относится и к генам, управляющим образованием желтого и зеленого пигментов (С1С2, С3, F1, F2, F3). Пока еще имеется мало доказательств в пользу такого заключения, но оно подтверждается работами по пигментообразованию у других организмов и, по-видимому, окажется справедливым и в данном случае; 3) различные цепи развития, контролируемые генами, взаимосвязаны. Образование пигмента начинается только на III стадии развития и тянется до V стадии. Если развитие структуры хлоропласта остановилось на III стадии, то хлоропласта могут вырабатывать желтые пигменты; могут ли они также вырабатывать хотя бы небольшое количество зеленого пигмента, зависит от характера дегенеративного изменения, которому они подверглись (мутация генов D или E). Вполне возможно, что существует также связь между двумя цепными процессами образования пигмента; 4) в нормальном развитии различные стадии взаимосвязаны, и каждая из них участвует в образовании конечного продукта - зеленого хлоропласта V.
Прежде чем расстаться с этим примером взаимодействия генов, следует указать, что хлоропластам свойственна определенная индивидуальность и что они не полностью находятся под управлением генов. Хлоропласт, подобно клетке, частью которой он является, образует новые хлоропласты путем деления, и клетка, потерявшая свои хлоропласты, не может заменить их. Иногда хлоропласт может утратить способность образования пигмента даже в клетке, содержащей все необходимые гены: гены готовы функционировать, но хлоропласт не может отвечать на это. Таким путем бесцветный хлоропласт дает начало клону бесцветных хлоропластов. Если это происходит на раннем этапе развития, весь побег становится бесцветным, и черенки, взятые с него, также будут бесцветными. Это явление заставляет помнить о том, что гены, хотя обязательны для жизни и развития, не обладают абсолютной монополией в управлении этим развитием. Мы вновь вернемся к этому вопросу в главе 28.
Гены, действующие серийно: сходство с работой на заводском конвейере. Гены, подобные A, B, D, G, или C1C2, C3, или F1, F2, F3 на рис. 78, действующие серийно, напоминают рабочих у конвейера; нефункционирующие гены этих серий являются как бы отсутствующими рабочими. Давайте посмотрим, что произойдет на конвейере линии, если один из рабочих отсутствует. На рис. 79 изображен сбор куклы четырьмя работницами у конвейера.
Перед Анной лежат головы кукол. Она присоединяет их к туловищам, поступающим к ней по конвейеру, и пересылает их к Бэрил. Эта девушка прикрепляет ручки и передает продукцию Кэти, которая пересылает продукцию к Дот, она одевает куклу и готовую укладывает в коробку. Допустим, что каждая работница может выполнять только свою собственную операцию и, более того, она может выполнять ее только тогда, когда все предшествующие манипуляции уже сделаны (например, в том случае, если различные части куклы прикрепляются друг к другу проволочками с крючками, которые можно использовать только в определенном порядке). Теперь представьте, что случится, если Анны не будет на ее месте. Если конвейер будет двигаться, безголовые, безногие и безрукие туловища будут проходить мимо места Анны, и так как другие работницы ничего не могут с ними сделать, то незаконченные куклы соберутся в конце конвейера. Если же на месте не будет Бэрил, Анна сможет выполнить свою операцию, а Кэти и Дот этого сделать не смогут, и куклы с головами, но без ног и рук соберутся в конце линии. Если отсутствует Кэти, соберутся безногие туловища; если не будет работать Дот, куклы получатся с головой и конечностями, но без одежды. Вообще, когда какой-нибудь рабочий отсутствует, на конвейере будет собираться недоделанная продукция, доставляемая предшествующим рабочим.
Предположим теперь, что мастер конвейера стремится выполнить план выпуска готовых кукол и имеет возможность получить готовых кукол или незаконченных кукол с параллельного конвейера, на котором избыток и тех, и других. Законченная продукция, конечно, выручит его, но, чтобы занять работой всех оставшихся работниц, он предпочтет взять незаконченную продукцию.
На рис. 79 видно, что это сделать невозможно, если отсутствует Дот, так как она является единственной работницей, одевающей кукол. Когда Кэти отсутствует, Дот может заниматься одеванием голых кукол, но Анна и Бэрил останутся без работы.
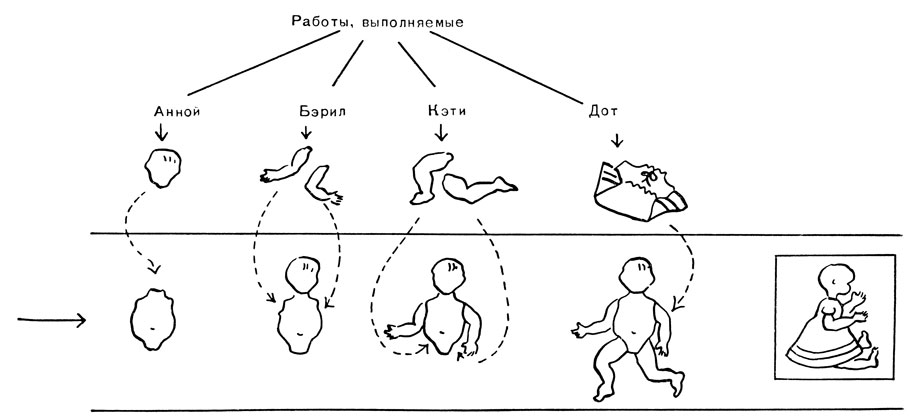
Рис. 79. Сбор куклы на конвейерной ленте
При отсутствии Бэрил девушка Дот опять-таки может быть занята одеванием кукол, но чтобы получить работу и для Кэти, мастер будет просить с другой линии кукол без ног, так как это обеспечит работой и Кэти, и Дот. И наконец, если отсутствует Анна, все остальные девушки могут быть заняты, если к Бэрил начнут поступать туловища кукол с головами, но без рук и ног. Если же этого не будет, то Кэти и Дот могут быть загружены работой, если будут получать кукол без ног, или только одна Дот будет занята одеванием кукол, поступающих к ней раздетыми, но полностью собранными. Таким образом, в целом предпочтительнее отсутствие работницы в начале конвейера; если отсутствует последняя работница, то мастера может выручить получение с другой линии только полностью оконченной продукции; если же отсутствует первая работница, то можно брать всю неоконченную продукцию с выполненной только первой операцией.
Разобрав этот пример, мы теперь сможем обсудить два способа серийного действия генов.
Синтез аминокислоты бактерией. Одной из основных составных частей живой материи является белок. Молекула белка представляет собой длинную цепь более простых органических молекул, называемых аминокислотами. Известно около 20 различных аминокислот, входящих в состав белков, и каждая из них, или почти каждая, необходима для жизни большинства живых организмов. Высшие животные, сходные с человеком, получают аминокислоты из белков пищи. Многие микроорганизмы, например большинство бактерий, "синтезируют" некоторые требуемые аминокислоты из простых неорганических составных частей их пищи. Всюду, где можно применить генетические методы для изучения синтеза аминокислот (в гл. 25 мы узнаем, что даже бактерии можно изучать генетически), обнаружили, что этот синтез идет по ступеням, из которых каждая управляется особым геном. Эти гены действуют серийно, как девушки на конвейере. Наш разбор "синтеза" куклы поможет понять, что происходит в том случае, когда один из генов мутирует в недеятельную форму.
На рис. 80 представлена обобщенная схема, которую можно применить ко многим отдельным случаям четырехступенчатого синтеза аминокислоты. Ген A, подобно Анне на рис. 79, осуществляет синтез первой ступени; ген B, как и Бэрил, - второй; ген C, как и Кэти, синтезирует третью ступень, и ген D, как и Дот,- четвертую. AA, так же как и одетая кукла, обозначает законченную молекулу аминокислоты; P1, P2 и P3 представляют собой три промежуточных продукта, или "предшественника", подобно трем незаконченным стадиям сбора куклы. Бактерии, у которых ген D не функционирует, могут выживать только в том случае, если им доставят извне готовую аминокислоту, так как ген D, подобно девушке Дот, является единственным геном, осуществляющим последнюю ступень синтеза. Бактерии с неактивным геном С могут существовать только при получении готовой аминокислоты или ее предшественника в виде P3. Бактерии с неактивным геном В могут использовать не только готовую аминокислоту или P3, но также и P2. И, наконец, бактерии с нефункционирующим геном A могут использовать как все три предшествующих продукта, так и готовую аминокислоту. Часто такие мутантные виды бактерий аккумулируют продукты, предшествующие месту разрыва в цепи синтеза: так, бактерии с неактивным геном B могут накапливать продукты P1 а бактерии с неактивным геном D - продукт P3.
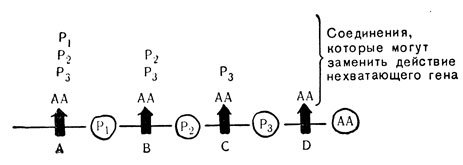
Рис. 80. Управляемый ступенчатый синтез одной аминокислоты: А, В, С, D - нормальные гены, управляющие стадиями синтеза; AA - аминокислот; P1, P2, P3 - промежуточных продуктов, предшественников AA; t - блокада серии реакций, в которой не действует один ген
Наука, изучающая химические реакции в живых организмах, называется биохимией. Изучение генов, управляющих биохимическими реакциями, называют биохимической генетикой.
Биохимическая генетика микроорганизмов открыла пути синтеза многих важных соединений, управляемых генами, действующими серийно. Имеется удивительное сходство между биохимическими путями синтеза таких важных соединений, как аминокислоты и витамины, клетками самых различных организмов, и биохимическая генетика микроорганизмов внесла существенный вклад как в биохимию, так и в генетику.
Биохимические ступенчатые реакции, управляемые генами, у людей. Познакомимся с примером из области генетики человека. В этом примере гены работают серийно, но не по синтезу, а по разрушению одной аминокислоты. В 1908 г. английский врач Гаррод опубликовал книгу под названием "Врожденные расстройства обмена веществ". В ней он описал много наследственных нарушений обмена веществ человека, т. е. тех биохимических процессов, с помощью которых живой организм синтезирует новые соединения или разрушает уже существующие. Одним из заболеваний, описанных Гарродом, была алкаптонурия - расстройство, характеризующееся потемнением мочи и потемнением и уплотнением хрящей. Заболевание наследуется по схеме рецессивного гена, лежащего в аутосоме, и, таким образом, должно быть обусловлено недействием одного единственного гена. При анализе мочи этих больных в ней нашли вещество, названное алкаптоном, в моче здоровых людей его нет. Теперь выяснили, что алкаптон является одним из продуктов распада белка или, вернее, аминокислоты, называемой фенилаланином. Когда больные получали избыточное количество этой аминокислоты с пищей, то содержание алкаптона в моче соответственно повышалось, чего не наблюдалось у здоровых людей. В процессе обмена веществ фенилаланин распадается вплоть до углекислого газа (CO2) и воды (H2O). По-видимому, алкаптон является промежуточным продуктом в цепи этого распада, и ген алкаптонурии прерывает цепь тотчас же после достижения стадии образования алкаптона. Мы делаем вывод, что нормальный аллель этого гена управляет ступенью, следующей тотчас же за образованием алкаптона.
В дальнейшем обнаружили, что второй ген, также аутосомный и рецессивный, вмешивается в ту же самую цепь обмена веществ реакций, но на более ранней ступени. В этом примере мутация, которая помогла открытию, приводит к чрезвычайно серьезным последствиям, так как гомозиготные люди страдают ярко выраженной формой слабоумия. Моча этих людей содержит большие количества вещества, называемого фенилпировиноградной кислотой, а само заболевание называют фенилпировиноградным слабоумием или фенилкетонурией (см. гл. 13). В процессе обмена фенил аланина фенилпировиноградная кислота является первым продуктом распада. Следовательно, у больных людей ее надо считать первой ступенью в цепи обменных реакций, которая блокируется мутантным геном; у здоровых людей этот первый этап распада осуществляется с помощью нормального аллеля. Весь описанный процесс представлен схематически на рис. 81.
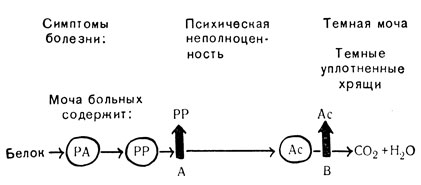
Рис. 81. Управляемые генами стадии расщепления аминокислоты (фенилаланина) у человека: А - нормальный ген, мутируя, дает фенилкетонурию; В - нормальный ген при мутации дает алкаптонурию; PA - фенилаланин; PP - фенилпировиноградная кислота; Ac - алкаитон
Заключение. Если теперь, после рассмотрения плейотропии взаимодействия генов, вернуться к рис. 72, станет ясно, что прямые линии, соединяющие гены и органы, должны быть заменены сетью промежуточных реакций, как показано на рис. 82. Однако и эта схема дает очень упрощенную картину исключительно многообразного и сложного взаимодействия реакций, управляемых генами, в процессе формирования данного фенотипа. Очевидно, что сложность сети, связывающей определенный признак с набором генов, будет отличаться у различных людей. Эта сеть гораздо более сложна, например, для размеров тела мыши, чем для присутствия данной аминокислоты в бактериальной клетке. Если мы спустимся в биохимическом анализе этой цепи процессов развития еще ниже, мы натолкнемся на вещество, являющееся непосредственным производным данного гена, связанным с ним одной-единственной ступенью. На рис. 82 это выражено одной линией, исходящей от каждого гена. Таким образом, плейотропия, которая, судя по рис. 72, как бы представляет собой свойство самих генов; здесь же она проявляется позднее как итог многочисленных связей межреакциями, управляемыми генами. Изучение веществ, создаваемых непосредственно генами,- один из животрепещущих вопросов современной генетики. Мы познакомимся с этим более подробно в следующей главе.
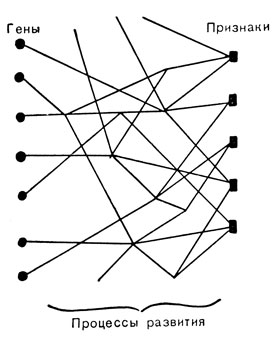
Рис. 82. Уточненная диаграмма генов и признаков
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'