22. История блестящего успеха
Гибридная кукуруза
Бывают моменты, когда необходимо немного вернуться назад, чтобы прийти к концу первым. В простом психологическом опыте животному и маленькому ребенку показывают привлекательный предмет, который можно получить, только обойдя вокруг загородки. Ребенок-ползунок быстро понимает, что ему надо повернуть в сторону от предмета, чтобы до него добраться; цыпленок этого никогда не понимает. Селекция растений многим обязана человеку, понявшему эту простую истину.
Под впечатлением экспериментов Иогансена с бобами Шелл в начале этого века сделал попытку вывести чистую линию кукурузы путем самооплодотворения. Кукуруза не относится к естественно самооплодотворяющимся растениям, как бобы, и она плохо переносит инбридинг. Поколение за поколением растения все мельчали и становились хилыми, а урожайность по зерну соответственно снижалась (рис. 69). С точки зрения практического выращивания кукурузы план Шелла казался совершенно нелепым, уводящим далеко в сторону от желаемой цели поднятия урожайности. Но Шелл не стремился к непосредственному практическому успеху; он хотел получить чистые линии для изучения генетики урожайности и был готов вернуться немного назад, на пути к тому, что в конечном итоге представляло собой шаг вперед.
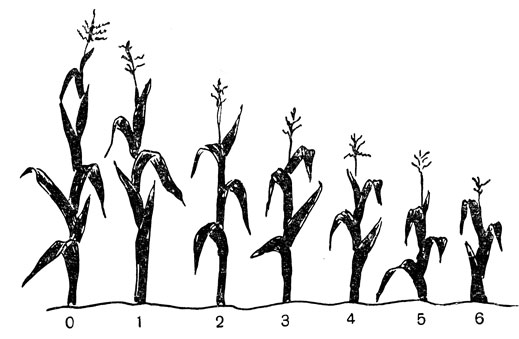
Рис. 69. Инбридная дегенерация у кукурузы
Как оказалось, реальным и важным шагом вперед в этой программе был не генетический анализ чистых линий, а наблюдение за тем, что скрещивание между слабыми инбридными линиями давало растения, превосходившие не только родительские экземпляры, но подчас и те сорта, от которых были выделены родительские линии (рис. 70).
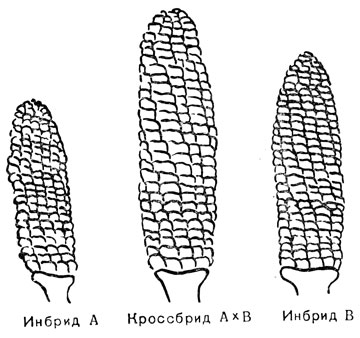
Рис. 70. Гибридная сила у кукурузы
Шелл предложил инбридные растения получать и сохранять с единственной целью использования их кроссбридного (гибридного) потомства.
Инбридное вырождение и гибридная сила были еще известны Ч. Дарвину, который провел много экспериментов по инбридингу и кроссбридингу. Некоторые селекционеры кукурузы в Соединенных Штатах предприняли работу по повышению урожайности кукурузы путем скрещивания инбридированных линий. Результаты превзошли все ожидания.
В начале 30-х годов гибридные семена впервые использовали в сельскохозяйственной практике. За время с 1935 по 1957 г. площадь земель, занятых гибридной кукурузой, в Соединенных Штатах возросла с 1,1 до 92,5%. В то же время общая площадь, занятая кукурузой, уменьшилась, так с каждого акра собирали все больше и больше зерна, так что 73000 акров в 1957 г. давали на 50% зерна больше, чем 100000 акров в 1930 г. В 1943 г. подсчитали, что повышение урожайности кукурузы вследствие использования гибридных семян составляло 669480000 бушелей1; при скармливании этой кукурузы ее было бы достаточно, чтобы обеспечить в год каждому мужчине, женщине и ребенку в стране по 24,5 кг мяса. В 1947 г. повышение урожая в результате использования гибридных семян составляло по крайней мере 800 млн. бушелей. Легко представить себе, что означало это колоссальное увеличение продовольственных продуктов в трудные годы войны и послевоенный период. После войны использование гибридной кукурузы распространилось во многие другие страны, например в Италию, Мексику, Россию. Некоторые страны, например Италия, импортировали американские гибридные семена; другие страны вывели собственные инбридные линии, от которых получили путем скрещивания гибридную кукурузу, приспособленную к местным условиям.
1 (1 бушель равен по емкости 36,3 л)
Гибридная кукуруза - пример выдающегося успеха современной генетики. Ученые, осуществлявшие план селекционной работы с кукурузой, работали, руководствуясь принципами Менделя. Попробуем проследить за ходом их мыслей и объяснить как вырождение при инбридинге, так и гибридную силу с точки зрения генетической науки. Самый важный факт, относящийся сюда, уже был упомянут в предыдущей главе. Было показано без объяснения, что "длительное самоопределение дает отдельные растения, гомозиготные по всем или почти по всем генам". Почему это так, легко понять из следующего простого примера.
Красная и слоновой кости окраска цветов львиного зева обусловлена парой аллеломорфов, ни один из которых не является полностью доминантным, поэтому гетерозиготные растения несут розовые цветы. В природе львиный зев опыляется пчелами, но этого можно избежать, надев "мешочки" на цветы и оградив их тем самым от посторонней пыльцы. На рис. 71 показано, что происходит при самоопылении гетерозиготного растения с розовыми цветами в его потомстве. В соответствии с первым законом Менделя (см. гл. 6) первое поколение расщепляется и дает 25% гомозиготных растений с красными цветками, 25% гомозиготных растений с цветками окраски слоновой кости и 50% гетерозиготных растений с розовыми цветками. Таким образом, первый акт самооплодотворения тотчас же дает 50% гомозигот по тому или иному аллелю. В следующем поколении, полученном при самоопылении, потомство от гомозиготных растении будет полностью гомозиготным, а потомство гетерозиготных растений вновь будет на 50% гомозиготным.
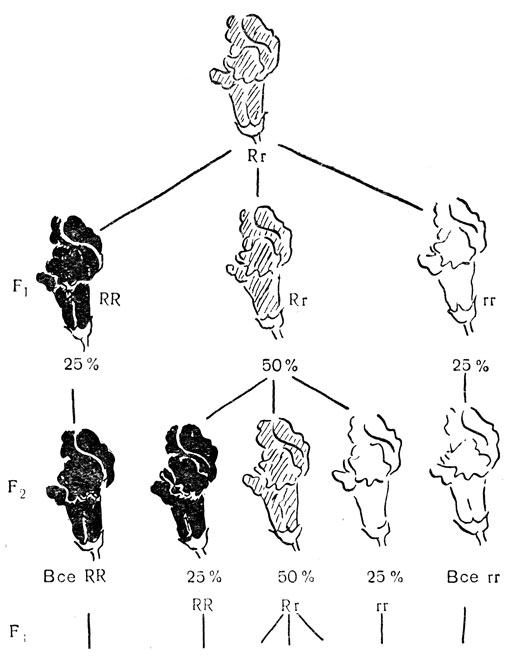
Рис. 71. Рост гомозиготности через самооплодотворение
Повышение гомозиготности при самооплодотворении. Этот процесс будет идти дальше, и оставшаяся часть гетерозиготных растений будет быстро уменьшаться с каждым поколением, пока, наконец, все гетерозиготные растения практически не исчезнут. Однако самым важным и заслуживающим запоминания является следующее: возникающие гомозиготы принадлежат к двум различным типам; Длительный инбридинг линии, начинающийся с гетерозиготы для одной пары аллелей, приводит в результате к двум гомозиготным линиям. Что справедливо в отношении одной пары аллелей, применимо и ко всем другим; длительный инбридинг превратит гетерозиготность по любому аллелю в линии, гомозиготные по тому или иному аллелю. Допустим, что мы начали действовать с растения, гетерозиготного по двум парам аллелей Aa и Bb; самооплодотворение приводит к линиям, гомозиготным по обеим парам. Теоретически возможно существование четырех таких линий, а именно, AABB, AAbb, aaBB, aabb. Если два гена лежат в различных хромосомах, все четыре линии будут получаться в приблизительно одинаковых численностях в соответствии со вторым законом Менделя (см. гл. 19). Даже если гены сцеплены (см. гл. 20), кроссинговер подчас будет допускать образование всех возможных линий, хотя и не в равных количествах. Чем больше аллелей, по которым исходное растение является гетерозиготным, тем больше гомозиготных линий может получиться при длительном самооплодотворении. Растения, опыляющиеся в природе перекрестно, обычно гетерозиготны по многим генам, и после длительного самооплодотворения каждое такое растение дает начало многим гомозиготным линиям.
Таким образом, выражаясь языком менделевской генетики, длительное самоопыление порождает набор отдельных линий, из них каждая является гомозиготной по комбинациям аллелей, по которым исходное растение было гетерозиготным. Остается спросить, почему это приводит к инбридному вырождению. Ответ уже намечался в гл. 12, где было показано, что даже умеренная степень инбридинга, наблюдаемая среди людей, может привести в определенных случаях к "дегенерации", например когда у двоюродных брата и сестры, гетерозиготных по гену амавротического слабоумия, родится гомозиготный больной ребенок. В общем же именно гомозиготность по прежде скрытым вредным рецессивам приводит к возникновению инбридных линий, худших по сравнению с родительскими. Итак, достаточно говорить об инбридинговом вырождении. Что можно сказать о гибридной силе, которая часто называется гетерозисом? Обобщая, можно сказать, что гибридная сила возникает в итоге восстановления гетерозиготности в процессе скрещивания. Мы уже познакомились с тем, что даже инбридные линии, возникшие первоначально из одного растения, отличаются комбинациями аллелей, по которым они являются гомозиготными; это применимо даже в большей мере к инбридным линиям, возникшим из различных сортов кукурузы. При этом почти каждая линия обязана своей дефективностью гомозиготности по различным рецессивным генам, и у гибридных экземпляров доминантные гены, внесенные одним из родителей, будут перекрывать рецессивные, полученные от другого родителя. Допустим, например, что a, b, c и d являются вредными рецессивами и что две инбридные линии имеют генотипы AAbbccdd и aaBBccDD. При скрещивании этих линий гибриды будут представлены генотипами AaBbccDd, т. е. они будут проявлять патологический дефект только одного из генов, в отличие от родителей, проявляющих патологический эффект по двум или трем генам. Очевидно, что степень, до которой гетерозиготность восстанавливается в итоге скрещивания, а также степень гетерозиса будет зависеть от выбора линий для скрещивания. В практике опытной проверке подвергали много различных скрещиваний, чтобы найти те линии, которые дают наиболее ценные гибриды.
Так как все растения инбридных линий не только одинаковы, но и гомозиготны, не происходит расщепления при образовании ими гамет, и все гибриды от двух инбридных линий характеризуются одинаковым генотипом. Гибриды начинают свой жизненный цикл с зерна на инбридном материнском растении, оплодотворенном пыльцой с иной инбридной линии. Такие зерна значительно более однородны по сравнению с зернами, собранными с обычных растений, и это иногда оказывается практически более ценным, например при заготовке консервов из сахарной кукурузы. С другой стороны, хилые инбридные растения дают мало зерен, вследствие этого рекомендуется пускать в хозяйственный посев гибридные семена, собранные с первого поколения гибридов, а не непосредственно с инбридных растений. Получающаяся в результате утеря однородности вследствие расщепления невелика, но если земледелец будет продолжать высевать семена с гибридных растений, он вскоре потеряет большую часть преимуществ гибридной кукурузы в результате расщепления; земледельцы, сеющие в своем хозяйстве гибридную кукурузу, должны покупать заново гибридные семена для каждого посева.
Хотя большинство генетиков придерживаются мнения, что инбридинг и гибридная сила в основном возникают описанным выше путем, некоторые генетики считают, что этим вопрос не исчерпывается. Они думают, что превосходство гетерозигот зависит не только от подавления вредных рецессивов, но также от наличия определенных пар аллелей, действующих лучше в гетерозиготной комбинации, а именно Aa превосходит как AA, так и aa. В главе 23 мы узнаем о паре аллелей человека, с которыми дело обстоит именно так. Вопрос о том, часто ли встречаются такие случаи и в какой степени они участвуют в формировании гибридной силы, в настоящее время вызывает многочисленные споры.
Планы использования гибридной силы в других областях растениеводства начали разрабатывать во многих странах. В племенном разведении домашних животных эти попытки встречают значительные трудности: невозможность самооплодотворения, резкое вырождение при инбридинге в результате длительных спариваний сестер и братьев, сниженная плодовитость и высокая стоимость отдельных животных. Нет никакой надежды достигнуть чего-нибудь и хотя бы приблизиться к гомозиготности по всем генам. Несмотря на это, даже скрещивание между частично инбридированными линиями позволяет проявиться гибридной силе, и обнадеживающие результаты были получены в птицеводстве. Выведение "гибридных цыплят" предпринял Г. Уоллес в 20-х годах, и теперь это дало блестящие результаты. Уоллес начал продавать первых гибридных цыплят в 1942 г., а в настоящее время продает ежегодно свыше 30 млн. Сходные работы проводят в свиноводстве.
Понятием, которому придается большое значение в литературе по гибридной силе, является "никинг" (удачная сочетаемость). Под этим подразумевают особую способность двух линий сочетаться при создании сильного гибрида; при этом дефективные гены одной линии погашаются их доминантными аллелями другой линии и наоборот. Мне хотелось бы закончить эту главу примером никинга из области генетики человека.
Глухонемота, как уже говорилось выше, часто определяется рецессивным геном. С точки зрения генетики глухонемые являются гомозиготными по этому гену, и когда два из них вступают в брак, надо ожидать, что все дети будут глухими. Между тем, к удивлению, в браках между глухими нередко появляются дети с нормальным слухом. Это происходит потому, что один или оба родителя в этих случаях стали глухими не от рождения, а приобрели глухоту в течение жизни из-за несчастного случая или болезни. Другие случаи объясняют по-иному.
Нормальный слух обусловлен правильным взаимодействием многих генов, и мутация в каком-нибудь из них может привести к глухоте. Рассмотрим два таких гена: D и E. Здоровые люди обладают генотипом DDEE, глухие генотипом ddEE или DDee. Кроме того, имеются гетерозиготы DdEE или DDEe или DdEe. Эти гетерозиготы слышат нормально, так как оба мутантных гена рецессивны по отношению к их нормальным аллеломорфам. В некоей семье передается мутантный ген d, и брак между двоюродными родственниками приводит к рождению глухой дочери Памелы. Генотип Памелы ddEE. Она воспитывается в приюте для глухих вместе со своим двоюродным братом Джеком, который, вероятнее всего, имеет тот же ген глухоты, что и Памела. Джек хочет жениться на Памеле, но, к счастью, она отказывает ему. Вряд ли можно сомневаться в том, что все дети от этого брака были бы генотипично ddEE и фенотипично глухими, как и их родители. Памела решает выйти замуж за другого воспитанника того же приюта Генри. Генри происходит из семьи, в которой передается ген e, его генотип DDee. В то время как гаметы Памелы несут гены d и E, гаметы Генри несут гены D и e. Генотип детей будет, таким образом, DdEe, и, к счастью семьи, все дети родятся с нормальным слухом. Таков простой пример никинга.
Читатель теперь сможет понять замечание, сделанное в главе 7 по поводу разведения норок. Там было сказано, что рецессивную платиновую мутацию получили независимо в двух норковых фермах, когда животных двух мутантных линий скрестили, то все потомство оказалось платинового цвета. Это дало генетикам основание считать, что обе мутации были одинаковы. Обоснование этого вывода теперь ясно. Если бы две мутации оказались различными, то потомство норок, так же как и дети Генри и Памелы, были бы гетерозиготны по обеим из них, и никинг подавил бы проявление каждого гена.
Никинг можно наблюдать во всех случаях, когда различные мутантные гены дают одинаковые фенотипические отклонения. Такие случаи встречаются очень часто; в чем здесь причина, будет разобрано в следующей главе.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'