25. Что представляет собой ген?
Хромосомы состоят из вещества, называемого нуклеопротеином, или ядерным белком. Нуклеопротеин состоит из двух компонентов: белка и нуклеиновой кислоты. Эта особая нуклеиновая кислота носит название дезокси-рибонуклеиновой кислоты, или просто ДНК. Другой вид нуклеиновой кислоты, называемый рибонуклеиновой кислотой, или РНК, обнаружен в цитоплазме, в некоторых вирусах и в небольшом количестве в хромосомах. До недавнего времени белок считали основным генетическим компонентом хромосом, главным образом потому, что он химически более сложен, чем нуклеиновая кислота, и поэтому казался веществом, подходящим для роли передатчика сложных распоряжений в цитоплазму. В частности, считали возможным допустить, что эти распоряжения передавались молекулами белка, которые, подобно молекулам антигена (см. рис. 22), свернуты в виде специфических образований. Ряд блестящих открытий, сделанных на протяжении последних 14 лет, заставил генетиков изменить свою точку зрения и считать ДНК главным компонентом гена. Ниже будет коротко сообщено о некоторых доводах, приведших к изменению этой точки зрения.
Мозаичная болезнь табака
На рис. 84 показаны три листа табака: левый лист взят со здорового растения, два других поражены так называемой мозаичной болезнью. Это очень заразное заболевание, быстро распространяющееся по посевам. В конце девятнадцатого века обнаружили, что здоровое растение табака можно заразить соком больного растения, даже если этот сок предварительно пропустить через фильтр из неглазированного фарфора, задерживающего все бактерии. Вскоре после этого обнаружили, что инфекционный возбудитель ящура также может проходить через фильтры, задерживающие бактерии. Такие возбудители инфекций получили название фильтрующихся вирусов; слово фильтрующиеся постепенно за ненадобностью отпало.

Рис. 84. Листья табака: Слева - нормальный в центре и справа - пораженные вирусом мозаики
До появления электронного микроскопа лишь самые крупные вирусы можно было наблюдать в виде мелких точек под самым большим увеличением. Теперь электронный микроскоп дает прекрасные картины даже мельчайших вирусов. На рис. 85, а представлен вирус мозаичной болезни табака под электронным микроскопом.
Химически вирусы состоят из нуклеопротеина. В некоторые вирусы, как и в гены высших организмов, в качестве нуклеиновых кислот входит ДНК, в других - РНК. Вирус мозаичной болезни табака принадлежит к последним. Молекула его белка имеет форму полого цилиндра, заключающегося в себе "палочку" РНК. На рис. 85, б показаны частички вируса, в которых химическим путем частично удален белок, так что конец палочки нуклеиновой кислоты выступает под ее оболочкой. Позже мы вернемся к этой картине.
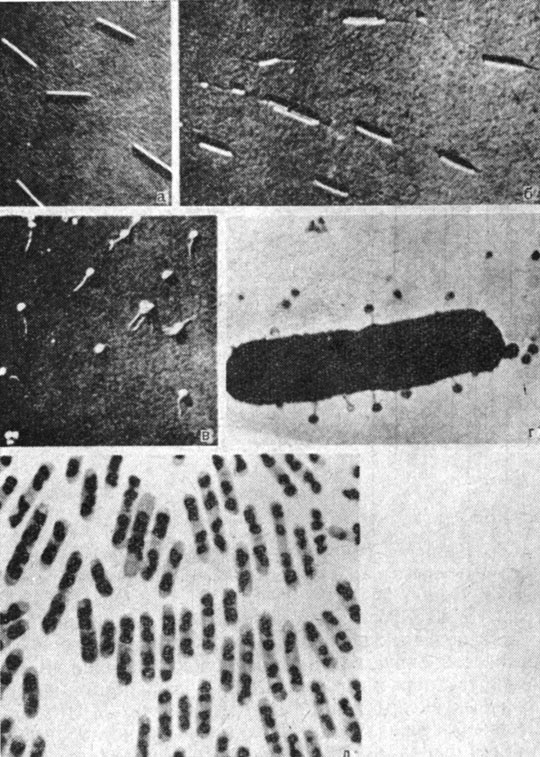
Рис. 85. Мозаичный вирус под микроскопом: а - частицы вируса табачной мозаики; б - частицы вируса табачной мозаики, отчасти освобожденные от белка; в - бактериофаги; г - бактериофаги, поражающие бактерию; д - бактерии с ядерными телами. Рис. а - г сделаны с фотографии под электронным микроскопом при увеличении в 10 - 15 раз, превышающем увеличение рис. д, полученного под обычным микроскопом при увеличении в несколько тысяч раз
Вирусы вызывают многие заболевания человека, животных, растений и даже бактерий. Они являются мельчайшими организмами (хотя вообще можно сомневаться в их праве называться организмами). Одним из свойств организма, даже паразитического, как, например, ленточного червя или дифтерийной палочки, является способность его к усвоению пищевых веществ. Вирусы утратили эту способность. Хотя они могут пребывать в живом состоянии вне клетки в течение длительного времени, особенно при низкой температуре, но в этих условиях не способны ни к усвоению веществ, ни к размножению. Они не могут это делать, даже находясь в самых сложных смесях питательных веществ, включая вытяжки из клеток, в которых они обычно паразитируют.
Настоящая жизнь вируса начинается только тогда, когда он попадает в живую клетку его хозяина. При этом вирус начинает вести себя как благоразумный завоеватель, который вместо разрушения сельского хозяйства и промышленности на захваченной у врага территории начинает использовать их в своих интересах. Клетка, сильно пораженная вирусом, использует собственные ферменты для размножения вирусов, подчас в больших количествах. Это приводит к гибели клетки и освобождению свежеобразовавшихся вирусных частиц, которые начинают внедряться и разрушать еще большее количество клеток.
В других случаях внедряющийся вирус ведет себя еще более благоразумно. Он оставляет клетке, в которой находится, достаточное количество питательных ресурсов для жизни и размножения и таким образом гарантирует себе сохранение собственной жизни и возможность размножения. При этом организм хозяина, например человека - носителя вирусного заболевания, давая пристанище вирусу, не обнаруживает внешних признаков его присутствия.
Вирус, находящийся внутри клетки хозяина, ведет себя очень сходно с геном. Ген, подобно вирусу, не может действовать или размножаться вне клетки. Такие выражения, как "ген воспроизводит самого себя" или "ген управляет синтезом белка", являются стенограммами, и их следует понимать так: "ген дает распоряжения клетке образовать другой ген, ему "подобный", или "синтезировать некий белок", так как весь строительный материал и нужные ферменты для этого синтеза содержатся в цитоплазме. В клетке, наводненной вирусом, распоряжения по образованию ферментов целиком или частично подаются вирусом и используются им для своего воспроизведения, а также для образования некоторых ферментов, необходимых для этой цели.
Сходство распространяется еще дальше. Наиболее поразительным фактом воспроизведения (репликации) генов является его точность. Клетка точно воспроизводит все гены, находящиеся в ней; если диплоидная клетка содержит два различных аллеломорфа определенного гена, она создает точную копию их обоих. Штаммы одного вируса, подобно аллеломорфам одного и того же гена, обнаруживают незначительное различие в своем действии; правый лист на рис. 84 несет одно блестящее желтое пятно, которое вызвано действием разновидности обычного вируса мозаичной болезни табака.
Такие разновидности возникают внезапно, подобно мутациям; и вновь возникающая разновидность вирусных частиц, подобно новым мутантным генам, реплицируется в точности внутри пораженной клетки. Иначе можно сказать, что каждый вариант штамма данного вируса, подобно каждому мутантному аллелю данного гена, отдает клетке свои собственные специфические приказания.
Как ген, так и вирус, состоят из нуклеопротеина. И по отношению к обоим возникает вопрос: подаются ли специфические приказания клетке посредством нуклеопротеина в целом или с помощью одного из его компонентов - белка или нуклеиновой кислоты? По отношению к гену возможны были пока только предположения, и они, как мы видели, склонялись в пользу того, что именно белок определяет специфичность гена. Применительно к вирусам оказалось возможным провести эксперименты, и они ясно показали, что носителем специфичности является нуклеиновая кислота, а не белок. В этих экспериментах использованы химические методы для отделения нуклеиновой кислоты от белка. На рис. 85, б показана промежуточная стадия этого процесса. Оказалось, что изолированная нуклеиновая кислота может заражать листья табака, а белок делать это неспособен. Более того, обнаружено, что нуклеиновая кислота подает все приказы для образования новых вирусов, так как заражение нуклеиновой кислотой, выделенной от определенного штамма, приводит к образованию вирусных частичек именно этого штамма. Наиболее поразительными оказались результаты опытов, в которых нуклеиновая кислота одного штамма смешивалась с белком другого. В этих смесях образовывались "восстановленные" вирусные частицы, сложившиеся из нуклеиновой кислоты одного штамма и покрытые белковой оболочкой из белка другого штамма. Заражение такими частицами всегда порождало новый вирус, который как по белку, так и по нуклеиновой кислоте оказывался принадлежащим к тому штамму, от которого была взята нуклеиновая кислота.
Таким образом, мы имеем ясный пример нуклеопротеина, в котором нуклеиновая кислота - и именно только нуклеиновая кислота - передает специфическую информацию клетке. Очевидно, было бы несколько поспешно предположить, что то же самое справедливо и для гена. Вирус мозаичной болезни табака и гены существенно различаются следующим: в то время как нуклеиновой кислотой гена является ДНК, нуклеиновой кислотой мозаичного вируса является РНК. Помимо этого, хотя и обнаружено, что сходство между вирусом и геном или группой генов очень велико, но еще не получено убедительных данных о том, что различия между штаммами мозаичного вируса табака сходны с различиями, возникающими под влиянием мутантных штампов. Ведь до сих пор у вируса мозаичной болезни табака не найдено ничего напоминающего половой процесс. Теперь обратимся к вирусам, у которых имеется половой процесс которые в качестве нуклеинового компонента содержат ДНК.
Бактериофаг
Бывают случаи в практике бактериологов, что культуры бактерий оказываются лизированными, т. е. бактерии как бы растворяются под влиянием чего-то возникающего внутри них. В начале нашего века открыли, что такой лизис бактерий вызывается вирусами, которые размножаются внутри бактерий и под конец разрывают их, освобождая большое количество новых вирусных частиц. Эти вирусы назвали бактериофагами (пожирателями бактерий); коротко их обычно называют фагами. Простой способ показать действие фагов заключается в распылении их по поверхности, целиком заросшей чувствительными бактериями. Каждая фаговая частица внедряется в бактерию. Когда бактерия лизируется, высвобождается много новых частиц, и они в свою очередь внедряются и лизируют соседние бактериальные клетки. В конце концов бактериальный "газон" оказывается разорванным круглыми дырами или "пятнами", каждое из которых указывает место, где фаг успешно недрился в бактерию.
Бактериофаги, как и другие вирусы, состоят из нуклеопротеина, но в противоположность вирусу табачной мозаики их нуклеиновая кислота представлена ДНК. Подобно вирусу табачной мозаики у них много сходного с генами. Бактериальная клетка, в которую внедрился бактериофаг, ведет себя так, как будто ей дана задача создать новый фаг, а не ген. Штаммы бактериофага, как и штаммы вируса табачной мозаики, различаются, например, по видам бактерий, в которых они с успехом внедряются, или по размерам пятен, которые они образуют. Как и у вируса табачной мозаики, эти различия обусловлены генетически, в том смысле, что новый вирус, созреваемый внутри бактерии, бывает точно такого же типа, как тот, который проник в бактерию. Как и у вируса табачной мозаики, штаммы с новыми свойствами могут возникать неожиданно, как бы вследствие мутации, и, возникнув однажды, сохраняют свой новый тип до тех пор, пока какая-либо другая мутация не изменит его.
Доказательство тому, что у бактериофага такие различия обусловлены мутациями истинных генов, получили при инфицировании бактерий смесью частиц от двух штаммов фага. Предположим, что штамм A дает крупные прозрачные пятна, а штамм B - мелкие, облакообразные и что большое количество бактерий заражено частицами обоих штаммов. Новые вирусные частицы, освобождающиеся из лизированных бактериальных клеток, при этом делятся на четыре типа: два первоначальных, образующих крупные прозрачные или маленькие облакообразные пятна, и два новых типа, образующих большие облакообразные или маленькие прозрачные пятна. Это напоминает результаты, полученные Менделем при скрещивании сортов гороха с круглыми желтыми и морщинистыми зелеными семенами (см. гл. 19). Описываемое явление показывает, что частицы фагов внутри клетки своего хозяина проходят какой-то процесс спаривания, в результате которого они обмениваются некоторыми из своих генов. На самом же деле сопоставление с данными Менделя не совсем удачно, так как гены, изученные Менделем, находились в различных парах хромосом и пересортировывались при мейозе, в то время как у бактериофагов мейоза нет и все гены находятся в одной и той же хромосоме. Новые комбинации генов различных фаговых частиц возникают в результате процесса, напоминающего кроссинговер. Исходя из этого, в нашем примере лучше провести параллель между четырьмя фаговыми типами и четырьмя типами сыновей, рождающихся от женщин, гетерозиготных как по гемофилии, так и по дальтонизму (см. гл. 20, рис. 62). В самом деле, правила сцепления и кроссинговера настолько хорошо применимы к генам бактериофага, что ни один хромосомный участок любого организма не имеет такой хорошей карты, как небольшой участок некоторых фаговых штаммов.
Фаговая генетика быстро развивается как раздел современной генетики, и не может быть сомнений в том, что бактериофаги несут истинные гены, расположенные в линейном порядке вдоль образования, которое можно назвать фаговой хромосомой. Образованы ли эти гены нуклеиновой кислотой или белком фаговой частицы?
Чисто химическое разделение фаговой частицы на ее компоненты невозможно без утраты способности к заражению, но сама частица подвергается грубому разделению каждый раз, когда она заражает бактерию. На рис. 85, в представлена фотография бактериофага под электронным микроскопом. Своей круглой формой и тонким хвостом он несколько напоминает головастика. Головка его содержит весь запас ДНК, хвост состоит полностью из белка. Когда бактериофаг атакует бактерию, он прикрепляется к добыче кончиком своего хвоста (см. рис. 85, г). Затем он проделывает отверстие в бактериальной стенке, используя хвост как инъекционную иглу, и впрыскивает содержимое головы в бактерию. Хотя впрыскиваемый материал главным образом состоит из нуклеиновой кислоты, но все же содержит небольшую примесь белка. Тем не менее эксперименты, описание которых может увести нас очень далеко от данной темы, показали, что гены бактериофага состоят из ДНК, а не из белка.
В итоге можно сказать, что исследования вируса табачной мозаики и бактериофага не дают полного ответа на вопрос: состоят ли гены из нуклеиновой кислоты или белка или же содержат оба вещества? На примере с вирусом мозаики ясно, что нуклеиновая кислота, и только она, передает клетке хозяина специфические указания по воспроизведению вирусов; но этой нуклеиновой кислотой является РНК, а не ДНК, и существование раздельных генов еще не доказано. ДНК бактериофага четко представлена в виде генов, которые по своим основным свойствам подобны генам высших организмов; но доказательство того, что эти гены состоят только из ДНК, пока еще косвенное. К счастью, существует особый тип частицы, давший неоспоримый ответ на обсуждаемый вопрос, по крайней мере для бактериальных генов. Эта частица называется "трансформирующим принципом", и мы его рассмотрим в следующей части. Одновременно это даст нам возможность коснуться особенностей половой жизни бактерии.
Трансформирующее начало - трансформация
Пневмонию вызывают бактерии, называемые пневмококками. Клетки этих бактерий имеют овальную форму и окружены капсулой. При введении пневмококков кроликам образуются антитела (см. гл. 10, рис. 22). Часть антител оказывает специфическое действие, направленное против капсул, а капсулы у различных штаммов различаются, поэтому антитела, выработанные против капсул одного штамма, реагируют с ограниченным числом капсул других штаммов. Это заставило бактериологов разделить пневмококки на типы в зависимости от их капсульных антигенов. Случается, что кокковая клетка теряет способность образовывать капсулу, и это передается всему ее потомству, что приводит к образованию клона бескапсульных клеток. Такие клоны утрачивают свою вирулентность, т. е. становятся безопасными для организма. В лабораторных условиях их легче всего распознать по тому факту, что на твердой среде они образуют колонии с шероховатой поверхностью в отличие от гладкой поверхности, образованной инкапсулированными бактериями. Поэтому стало обычным употребление терминов "шероховатый" и "гладкий" по отношению к пневмококкам без капсулы и с капсулой.
Шероховатые штаммы нельзя разбивать на типы, так как они не вызывают образования капсульных антител при введении животным. Однако изредка бактерия шероховатого штамма вновь приобретает способность к образованию капсулы; тогда она дает начало гладкому клону того самого типа, что и клон, из которого образовался "шероховатый" клон вследствие потери способности к образованию капсулы. Таким образом, шероховатые штаммы возвращаются к типу гладкого штамма, из которого они когда-то возникли.
В 1928 г. один английский бактериолог провел замечательный эксперимент. Он заразил мышей смесью из бактерий шероховатого штамма и гладкого штамма, но убитого при нагревании. В отдельности ни один из компонентов смеси штаммов не был вреден для животных. Смесью же был убит значительный процент мышей. Оказалось, что соединение живых бактерий шероховатых штаммов с убитыми бактериями гладкого штамма каким-то образом восстанавливало вирулентность первых; действительно, при исследовании бактерии погибших мышей обнаружили, что они имели капсулу и образовывали гладкие колонии. Наиболее поразительное явление в этих экспериментах наблюдалось, когда шероховатые и гладкие бактерии в первоначальной смеси принадлежали к различным типам; в этих случаях шероховатые бактерии трансформировались в гладкие того типа, к которому принадлежали убитые гладкие формы. Если, например, шероховатые бактерии принадлежали к штамму II типа, а убитые гладкие формы - к штамму III типа, то шероховатые бактерии трансформировались в гладкие III типа. То, что это явление не сводилось к простому окружению шероховатых бактерий капсулами убитых гладких форм, следовало из факта, что все потомки оказывались принадлежащими к III типу. Во всех подобных экспериментах убитые гладкие бактерии трансформировали шероховатые в их собственный капсульный тип.
Эти опыты в свое время не привлекли должного внимания. Почти 20 лет спустя такую же работу провели американские ученые. Они обнаружили, что трансформация не зависела от присутствия убитых бактерий и могла быть проведена с бесклеточной вытяжкой из гладких бактерий. Когда эти вытяжки фракционировали на составляющие их химические вещества, выяснилось, что вся трансформационная способность заключалась в ДНК, которая поэтому была названа "трансформирующим началом". Как только это установили, трансформацию стали наблюдать у других видов бактерий и для других наследственных свойств. Так, чувствительные к антибиотикам - пенициллину или стрептомицину - бактериальные штаммы могут приобретать стойкую устойчивость к антибиотикам путем трансформации с помощью ДНК, выделенной из стойких к антибиотикам штаммов.
Таким образом, трансформация представляет собой передачу наследственных свойств от одной клетки к другой с помощью чистой ДНК. Были выдвинуты различные гипотезы для объяснения этого поистине изумительного процесса. Сторонники одной из них предположили, что трансформирующее начало представляет собой изолированный ген или группу нескольких генов, которые каким-то образом не только проникают в живую бактериальную клетку, но также и прочно внедряются в генетическую субстанцию своего хозяина, и, следовательно, трансформацию можно рассматривать как трансплантацию гена от одной особи к другой. В последующем была доказана правильность этой гипотезы, но это оказалось возможным после более глубокого изучения генетики бактерий.
На рис. 85, д показаны бактерии, окрашенные одним из красителей, применяемых для окраски ядер высших организмов. Можно видеть, что каждая клетка содержит несколько окрашенных тел, так называемых ядерных тел. Их структура проще по сравнению с истинными ядрами; например, они не окружены мембраной подобно ядру. Тем не менее установлено, что в них помещается генетическое вещество клетки.
Доказательство тому, что это вещество состоит из генов, расположенных вдоль одной хромосомы, было получено в генетических опытах, впервые поставленных Ледербергером и Татумом, которые в 1959 г. получили Нобелевскую премию за открытие генетических рекомбинаций у бактерий. Эти ученые обнаружили, что смесь бактерий с различными наследственными свойствами может дать рост штаммов, которые сочетают свойства одного штамма с иным свойством второго штамма. Мы знаем, что это явление считается основным признаком полового процесса. У бактерий обнаружение половых процессов при появлении новых "рекомбинантных" типов может быть значительно облегчено использованием так называемых методов отсеивания. Эти методы основаны на том, что с их помощью убиваются все бактерии, за исключением небольшого количества таких, которые путем половых рекомбинаций стали обладателями двух свойств, имевшихся в отдельности у двух родительских штаммов. В качестве примера мы можем рассмотреть два штамма кишечных бактерий A и B. Штамм A может расти на галактозе (см. гл. 24), но погибает от стрептомицина. Штамм B резистентен к стрептомицину, но не может расти на галактозе. Если большое количество бактерий обоих штаммов поместить вместе в среду, содержащую стрептомицин и галактозу без других Сахаров, клетки штамма A погибнут от стрептомицина, а клетки B - от голода. Выживут только клетки, которые будут сочетать способность расти на галактозе, унаследованную от штамма A, с устойчивостью к стрептомицину, унаследованную от штамма B.
До настоящего времени "спаривание" наблюдали только у некоторых штаммов бактерий. Даже у них поначалу казалось, что спаривание происходит очень редко, но эта точка зрения изменилась, когда открытие двух полов у бактерий сделало возможным смешивать мужские и женские клетки в соответствующих соотношениях. Различие между полами у бактерий сходно с различиями у высших организмов, у которых представители женского пола являются пассивными партнерами, а мужского - активными. Когда бактериальные клетки спариваются, они соприкасаются друг с другом - и генетическое вещество переходит от мужского партнера к женскому. В этом явлении процесс сов" падает в общих чертах с половым размножением. Различие только в способе передачи генов от мужского партнера женскому. Это происходит путем выталкивания его хромосомы в продольном направлении в женскую клетку со столь закономерной скоростью, что можно предсказать, через сколько минут после первого контакта между мужской и женской клетками данный ген перейдет в женскую. Видимо, две клетки расходятся прежде, чем вся мужская хромосома успеет перейти в женскую клетку, так как обычно женская клетка после спаривания содержит только часть мужской хромосомы в добавление к собственной. Если гены мужской хромосомы отличаются от соответствующих генов женской, то кроссинговер приведет к новым комбинациям генов.
Бактерии, которые не скрещиваются описанным способом, могут применять другие, даже более оригинальные способы обмена генов и создания новых генных комбинаций. Помимо того, что в природе может происходить трансформация живых клеток случайно убитыми, многие бактерии отдают один или несколько своих генов частицам фагов, которые потом переносят их в новую клетку, где они вселяются в хромосому.
Исследования этих видов половых процессов у бактерий установили существование бактериальных генов. Они показали, что трансформация состоит в передаче генов от умерших клеток живым и внедрении их в хромосому-хозяина.
Таким образом, мы убедились: трансформирующим началом является ДНК, и вопрос о том, что бактериальные гены состоят из ДНК, решен. Нет оснований сомневаться в том, что и у высших организмов ДНК также образует основную, возможно, самую важную часть гена. Но роль белка, который тесно связан с ДНК в хромосомах всех высших организмов, остается невыясненной. Между тем исследования в области физической химии показали, как приспособлена структура ДНК к тем функциям, которые должны выполнять гены.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'