26. Дезоксирибонуклеиновая кислота
ДНК - дезоксирибонуклеиновая кислота, подобно белку, образует гигантские молекулы, состоящие из более мелких и более простых слагаемых - кирпичиков. Кирпичики белка носят название аминокислот (см. гл. 23). Кирпичики, слагающие нуклеиновые кислоты, называются нуклеотидами. В свою очередь нуклеотид состоит из трех частей: молекулы сахара, фосфатной группы и так называемого пуринового или пиримидинового основания. Пурины и пиримидины представляют собой органические молекулы типа встречающихся часто во многих природных продуктах.
Так, например, к пуринам относится кофеин. Все нуклеотиды ДНК содержат одну и ту же молекулу сахара, называемую дезоксирибозой; нуклеотиды РНК, т. е. рибонуклеиновой кислоты, содержат сахар, называемый рибозой, отличающийся от дезоксирибозы тем, что в нем содержится дополнительный атом кислорода.
Основания в различных нуклеотидах ДНК не все одинаковы; имеется четыре основания: два пурина и два пиримидина, и каждый нуклеотид несет одно из них. В разных случаях может встречаться и пятое основание. Мы углубились бы слишком далеко в химию, если бы пожелали приводить здесь формулы пуринов и пиримидинов ДНК; но изложение в последующем будет облегчено, если мы узнаем, как они называются. Два пурина называются аденином (A) и гуанином (G); два пиримидина - тимином (T) и цитозином (C). Вначале думали, что гигантская молекула ДНК всегда содержит одинаковое число A, G, T и C; фактически каждую такую гигантскую молекулу можно подразделить на группы из четырех нуклеотидов, несущие в своем составе по одной A, G, T и C. Если бы это было правильно, то возможности ДНК как переносчика генетического кода были бы довольно ограниченными; как легко можно проверить, каждая группа из четырех нуклеотидов в лучшем случае могла бы дать лишь 2×3×4=24 различные "буквы" и понадобилось бы очень много таких групп по четыре для написания даже простого "слова", хотя для развития даже самого простого организма нужно иметь тысячи "слов", т. е. процессов, управляемых генами. До недавнего времени эти соображения склоняли большинство генетиков к рассмотрению белковой части хромосомы как существенного переносчика генетической информации.
Только что описанное представление о строении ДНК сейчас оставлено. Соотношение между четырьмя основаниями в молекуле ДНК различно у разных видов и не всегда 1:1:1:1. Нет больше никаких доводов принимать, что основания в молекулах ДНК составлены группами по четыре. Весьма вероятно, они лежат в любой возможной последовательности, вроде AATG G GA... или TG TG CCA... и так далее. Это сразу позволяет нам избавиться от ограничений в отношении кодирования. Очевидно, что даже очень короткие последовательности трех соседних нуклеотидов могут "произнести" много разнообразных "букв", если допустить всевозможные комбинации и перестановки из четырех оснований.
Три нуклеотида, например, можно записать как AAA, или CCC, или AGA и так далее. Для бактерий и вирусов, где ДНК несет генетическую информацию, разумно принять, что код записывается последовательностью нуклеотидов; резонно допустить, что это приложимо и к более высокоорганизованным формам.
Сохраняется, однако, своеобразное числовое соотношение для четырех оснований в ДНК: число пуринов всегда равно числу пиримидинов (A+G=T+C). Более пристальный анализ показывает, что прилаженность пуринов и пиримидинов идет еще дальше и число пуринов оказывается равным числу тиминов (A=T), тогда как число гуанинов оказывается равным числу цитозинов (G=C). Например, если в ДНК данного вида организмов имеется превышение количества аденина над гуанином, обнаруживается и соответственное превышение тимина над цитозином. Это химическое свойство ДНК оставалось непонятным до 1953 г., когда в Кембридже два молодых ученых англичанин Крик и американец Уотсон выдвинули новую идею о строении ДНК. Их схема не только объяснила любопытное количественное соотношение между основаниями; она позволила просто истолковать наиболее интригующее биологическое свойство генетического материла, а именно его способность к репликации. Прежде чем разобраться в уотсон-криковской м ли ДНК, надо познакомиться с трудностями, которые стояли перед генетиками при попытках понять репликацию генов.
Трудно представить себе сходные процессы в природе, которые позволили бы дать ключ к раскрытию этой тайны. Когда кристалл, например, обычной поваренной соли держат в крепком растворе этой соли, он растет, и новые молекулы соли добавляются таким образом, что характерная форма кристалла, в данном случае куб, неизменно сохраняется.
Получающийся большой кристалл легко может быть опять расщеплен на более мелкие кубики. Имеется некоторое сходство между репликацией генов и процессом роста и подразделения по строго определенной форме; однако различия более разительны, чем черты сходства. Кристалл обладает простой молекулярной структурой; его атомы лежат в строгом порядке. Гены, какова бы ни была их особая химическая структура, несут столь высоко специализированную функцию, что должны отличаться друг от друга весьма тонким и специфическим образом. Эти свойства тонкости четко репродуцируются при размножении генов, репродукция не может происходить под действием простых физических сил, вроде тех, которые управляют ростом кристаллов. Гораздо лучше это сопоставить с образованием антител. Антигены (см. гл. 10) походят на гены своей высокой степенью специфичности, и они переносят эту специфичность на антитела, образование которых они вызывают. Гипотеза о том, как клетка изготовляет копию генов, близка к гипотезе о копировании антителом специфичности антигена и очень привлекательна, однако для ее принятия имеются некоторые трудности. Антитело не является копией антигена; их отношение друг к другу таково, как отношение медали и формы, в которую наливают металл (см. рис. 22). Чтобы воспроизвести саму антигенную молекулу, антитело должно было бы служить формой для изготовления антигена, который в самом деле должен быть абсолютно схож с исходным.
Таким образом, теории воспроизводства генов, сопоставляющие это явление с образованием антител, должны признавать, что ген - форма для каких-то промежуточных продуктов, которые, в свою очередь, будут формой для новых генов. Выдающейся заслугой Уотсона и Крика надо считать то, что они объясняют образование генной репликации без обращения к гипотетическому промежуточному носителю специфичности.
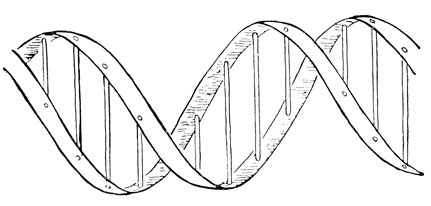
Рис. 86. Структура ДНК по Уотсону и Крику
Уотсон и Крик в основу своей схемы положили картину дифракции ДНК в рентгеновских лучах. Когда лучи Рентгена проходят через кристалл, они рассеиваются атомами, и, смотря на рисунок, образуемый этими лучами на рентгеновской пластинке, ученые могут делать выводы об относительном расположении атомов. Эту технику применили к веществам, не являющимся истинными кристаллами, но молекулы которых, подобно белку и нуклеиновой кислоте, состоят из составных частей, правильно расположенных на определенном расстоянии. Дифракционные исследования по нуклеиновой кислоте выполнялись на протяжении многих лет в Лондоне и Кембридже. В 1953 г. Уотсон и Крик предложили по-новому осмысливать эти данные (рис. 86). Молекула ДНК показана частично; она состоит из двух спирально завитых вокруг друг друга нитей. Поскольку диаметр завитков остается тем же на протяжении всей молекулы, более правильно говорить об улиткообразном завивании и называть всю структуру двойной улиткой. В действительности, конечно, ленты не представляют собой плотных волокон, но состоят из отдельных атомов, удерживаемых при помощи сильных химических связей. Более слабые связи соединяют ленты накрест и удерживают их вместе. В химическом отношении ленты состоят из альтернирующих (чередующихся) фосфатных и сахарных единиц; основания привязываются к сахару, одно основание к каждой сахарной единице, и выступают в полый цилиндр, образуемый двумя завивающимися лентами. Противоположные основания удерживаются слабыми химическими связями.
Если представить себе, что ленты развились и вся структура стала плоской, то она выглядела бы примерно так:
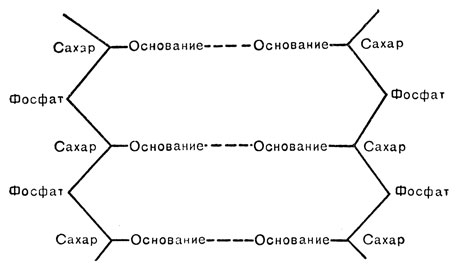
Надо теперь сказать, что из двух типов оснований пурины более крупные, чем пиримидины. Если бы пиримидин одной ленты попарно соединялся с пиримидином другой, диаметр всей структуры в данном участке был бы меньше, чем в участке, где попарно соединяются два пурина. Если желательно сохранить одну и ту же толщину на всем протяжении молекулы ДНК, то надо расположить основания таким образом, чтобы на каждом звене пурин приходился против пиримидина. Так как дифракционная рентгеновская картина показывает, что диаметр ДНК не меняется от участка к участку, основания и должны на самом деле так располагаться. Вам становится понятным, что такое предположение автоматически приводит к равенству соотношений пуринов и пиримидинов и объясняет до сих пор загадочное численное соотношение между основаниями. Более того, если принять во внимание химическую структуру отдельных оснований, выявляется, что, для того чтобы уложиться в имеющееся пространство, аденин должен идти в паре с тимином, а гуанин - с цитозином. Становится понятным, что в молекуле ДНК всегда находится столько же аденинов, сколько тиминов, и столько гуанинов - сколько цитозинов. Последовательность основания в двух лентах как бы дополняет друг друга: если мы берем произвольно одно из них, то другое уже не может быть любым. Так, например, если в каком-то участке ленты последовательность такова ...AGGTTCTGAC..., то против нее неизбежна последовательность TCCAAGACTG... Надо остановиться еще на огромной привлекательности модели Уотсона - Крика для генетиков: она состоит в предположении о простоте механизма репликации генов.
Согласно современной теории, ДНК реплицируется следующим образом: во-первых, обе завитые ленты развиваются и отделяются; как это происходит, до сих пор не вполне ясно. Как только ленты становятся свободными, основания, лежащие последовательно, оказываются окруженными жидкостью, в которой содержатся разнообразные химические строительные кирпичики, среди них фосфаты, сахар и четыре пуриновых и пиримидиновых основания, необходимые для синтеза ДНК. Из этого запаса строительного материала каждое основание исходной ленты привязывает к себе новое основание, и эти новые основания затем связываются друг с другом новой сахаро-фосфатной лентой, которая начинает обвиваться вокруг старой и образовывать с ней двойную улиткообразную структуру. Наиболее важным обстоятельством всего этого процесса надо считать то, что старые основания не свободны в выборе новых. Если каждое основание не будет подбирать свое дополнительное к нему, иными словами, если A не будет выбирать T, T - A, G - C и C - G, в новой "улитке" образуется утончение там, где произойдет неверный выбор, и это приведет к тому, что непригодное основание будет замещено подходящим. На диаграмме ниже жирным шрифтом отмечены новые основания и ленты.
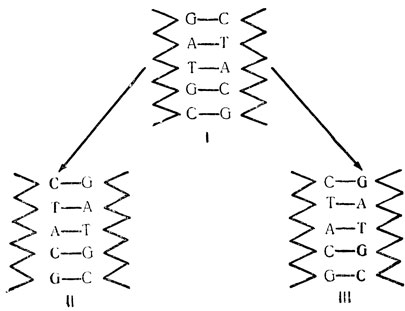
На этой диаграмме показаны основные черты репликации ДНК согласно современной теории: каждая дочерняя хромосома образуется из одной старой ленты и одной новой, и специфическая последовательность в двух дочерних хромосомах (II и III) такая же, как в исходной (I).
Подобно любому крупному открытию, модель Уотсона - Крика выдвинула больше вопросов, чем смогла дать ответов. Вопросы эти таковы: используется ли для кодирования вся ДНК в хромосоме или в хромосоме лежат "не имеющие смысла" участки, играющие роль знаков препинания - точек или запятых или служащие для каких-то совершенно иных целей. Привязана ли генетическая информация только ДНК даже в высших организмах, в которых хромосомная ДНК всегда тесно связана с белком и некоторым количеством РНК? Как прочитывается этот код и переводится в специальные биохимические процессы? РНК, очевидно, играет ведущую роль в этом деле; как же именно это осуществляется?
Поставив эти вопросы, мы подошли вплотную к рубежам науки сегодняшнего дня. Углубляться в них - это значит выходить за пределы задач, поставленных в этой книге. Однако в следующей главе мы увидим, как открытие Уотсона и Крика помогло нашим попыткам ответить на старый вопрос: что же представляют собой мутации и как они возникают?
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'