28. Ведущая роль генов в наследственности
До сих пор мы рассматривали лишь случаи, в которых передача признака зависит от генов, хотя проявление признака может зависеть и от влияния среды. Так как гены заключены в ядра, такой тип наследственности носит название ядерной. Подавляющее большинство хорошо изученных случаев наследственности относится к ядерному типу, но это не исключает возможности внеядерной, или цитоплазматической, наследственности. С одним примером внеядерной наследственности мы уже познакомились в главе 23. Как было показано, образование хлорофилла находится под контролем большого числа генов; однако эти гены требуют соучастия хлоропластов. В хлоропластах могут возникать изменения, которые делают их неспособными реагировать на гены-регуляторы, и эти изменения передаются от первого, ставшего ненормальным хлоропласта ко всем его потомкам. Известно много примеров хлоропластной наследственности, и, так как хлоропласты размножаются, подобно генам, путем репликации и деления, этот тип внеядерной наследственности довольно хорошо изучен.
Имеются другие случаи хлоропластной наследственности, и их можно обнаружить с помощью специальных методов. Имеется два правила их обнаружения: одно отрицательное, а другое положительное. Первое правило гласит, что наследственные черты, передаваемые через цитоплазму, не дают явления менделевского расщепления у гибридов. Положительное правило основывается на том, что у всех высших организмов в женской гамете много цитоплазмы, а в мужской ее очень мало. Можно поэтому ожидать, что признак, передаваемый цитоплазматически, будет наследоваться по материнской линии. Это оказывается действительным для хлоропластной наследственности. При скрещивании растений с нормальными и ненормальными хлоропластами все потомки оказываются с нормальными хлоропластами, если материнское растение имело нормальные хлоропласты, и все потомки оказываются с ненормальными хлоропластами, если в материнском растении находились ненормальные хлоропласты. Изредка хлоропласты могут передаться потомству и через пыльцу. Это, однако, не меняет существенность разницы между потомством обоих реципрокных (обратных) скрещиваний.
Сопоставление результатов реципрокных скрещиваний может считаться полезным приемом в поисках цитоплазматической наследственности. Этим приемом надо, однако, пользоваться с осторожностью, потому что разница между потомствами от перекрестных скрещиваний может появляться, если наследственность идет чисто ядерным путем, самым простым примером может служить связанная с полом наследственность (см. гл. 15). Скрещивание золотистого петуха ss и серебристой курицы S дает в потомстве серебристых Ss петушков и золотистых курочек s (см. рис. 42); обратное (реципрокное) скрещивание серебристого петуха SS и золотистой курицы s дает только серебристое потомство: петушки несут факторы Ss, а курочки - S. Такие случаи легко отличимы от цитоплазматической наследственности.
Другая причина отличий в реципрокных скрещиваниях - это раннее проявление генов в неоплодотворенном яйце. Такое явление показано на рис. 89. У мучной моли Ephestia нормальный черный цвет глаз у гусениц и бабочек во время развития проходит через ряд биохимических стадий (см. гл. 23, рис. 80, 81). Одна из этих стадий выходит из строя, если ген A заменить его аллеломорфом a. Ген A доминирует над a, и глаза у гусениц и взрослых молей Aa оказываются черными, такими, как у гомозигот AA. Глаза у aa насекомых светлые, так как одно из необходимых веществ-предшественников не образуется в отсутствие A. Если скрестить гетерозиготного черноглазого самца Aa со светлоглазой самкой aa (см. рис. 89, I), потомство расщепляется на 50% Aa гусениц с черными глазами и 50% aa гусениц со светлыми глазами в полном согласии с тем, что можно предсказать, исходя из первого закона Менделя (см. рис. 51).
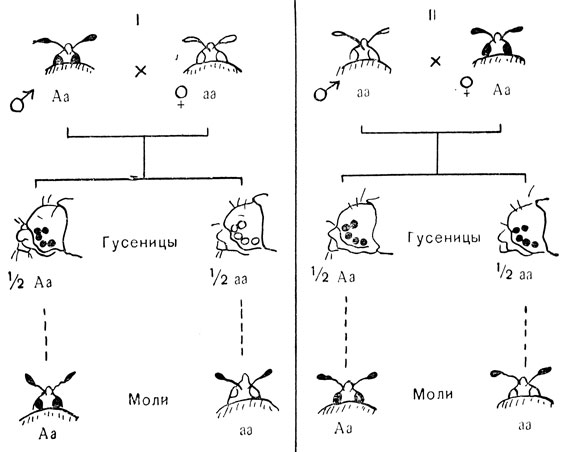
Рис. 89. Материнская наследственность у мучной моли Ephestia: А - ген темных глаз; а - ген светлых глаз
Обратное скрещивание (см. рис. 89, II) дает другой результат. У всех гусениц оказываются черные глаза. При этом все же получается расщепление на Aa и aa особи, что можно наблюдать, следя за молями, когда они выходят из коконов: у половины из них глаза черные, а у другой - светлые, совершенно сходно с тем, что наблюдается в скрещивании I. По-видимому, ген A матери вырабатывает в яйце достаточное количество вещества-предшественника, и его хватает на всю жизнь насекомого в стадии гусеницы; когда этот запас исчезает, больше вещества в отсутствие гена A образоваться не может, и в глазу (окрыленной) моли не образуется черного пигмента. Опять становится ясно, что здесь мы имеет дело с ядерной, а не цитоплазматической наследственностью. Разделение типов наследственности было бы не столь простым, если бы действие материнского гена распространялось на всю жизнь потомства. Это может также привести к случаям, которые на первый взгляд напоминают цитоплазматическую наследственность. Рис. 90, II. изображает два типа завивания раковины у пресноводного моллюска Limnea. В индивидуальном развитии левизна и правизна завитков раковины зависят от расположения первых четырех клеток молодого эмбриона (см. рис. 90, II). Это, в свою очередь, зависит от асимметричной структуры, заложенной в неоплодотворенном яйце под влиянием генов матери. Этой асимметрией управляет пара генов. Ген для правой асимметрии D доминирует над своим рецессивным аллелем c, дающим левую спиральность. Материнская особь генотипа DD или Dd дает вне зависимости от генотипа отца потомство с правой спиральностью. Сходным образом самка генотипа dd дает только левозавитое потомство. В общем, направление спирали любой особи определяется генотипом ее матери, но не ее собственным генотипом. Это легко проследить на схеме разведения, изображенной на рис. 91. При составлении этой схемы было принято во внимание, что улитки являются гермафродитами. Это значит, что у них имеются одновременно и мужские, и женские половые органы и они способны как к самооплодотворению, так и к перекрестному оплодотворению. Если бы опыты скрещивания ограничились первым поколением, создалось бы впечатление, что перед нами случай четкой цитоплазматической наследственности (согласно установленным нами закономерностям). Так как не было расщепления и в обратных скрещиваниях, все потомки обладали материнским фенотипом.
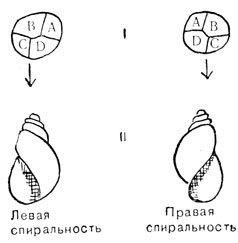
Рис. 90. Правое и левое завивание раковины прудовой улитки: I - первые четыре клетки развивающегося эмбриона. II - сформированные раковины
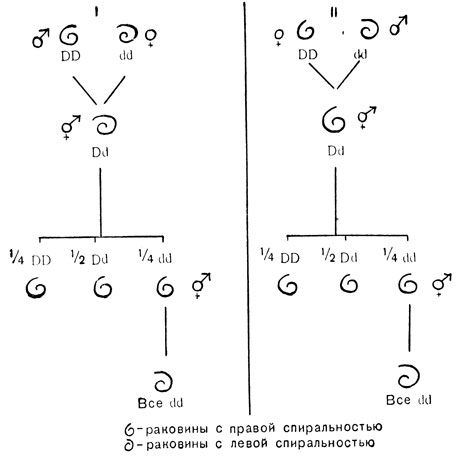
Рис. 91. Материнская наследственность у Limnea: d - рецессивная аллель для левой спиральности; D - то же для правой спиральности; ♂ - использован в скрещивании как ♂; ♀ - использована в скрещивании как ♀ ♂ - самоосеменяющееся животное
Если оставить в стороне все сходные случаи материнской наследственности ядерного действия, останутся примеры, которые действительно должны быть отнесены к цитоплазматической наследственности или по крайней мере к наследованию через цитоплазму. Эти выражения нельзя считать чисто словесными отличиями; имеются опыты, говорящие о том, что ряд случаев, казавшихся примерами настоящей цитоплазматической наследственности, мог быть объяснен передачей микроорганизмов через цитоплазму. Наиболее показательным примером надо считать явление "убийц" и "убийц половых партнеров" у микроскопического пресноводного животного Paramecium (рис. 92). Этот организм представлен одной клеткой и дает клоны путем непрерывного деления на две клетки; вдобавок к этому у них наблюдается скрещивание и менделевское расщепление генов. Когда разные клоны парамеций оказываются в одной банке, случается, что особи одного клона убивают выделяемым секретом особи другого клона. Для самих убийц этот секрет безвреден. Клоны различных убийц обладают различными методами убивания своих жертв. Большинство из них не убивает своих партнеров спаривания; однако имеются штаммы, которые вместо убивания на расстоянии при помощи секрета убивают своих партнеров (в процессе конъюгации) через тесный контакт. Свойство убийцы заложено как в ядре, так и в протоплазме. Явление весьма сходному обычных "убийц" и "убийц партнеров конъюгации". Описание дается для первых. Ядерная основа явления состоит в наличии пары аллелей К и к. Животные, гомозиготные по фактору к, оказываются чувствительными к убивающему воздействию, и сами не могут сделаться убийцами:, Животные, несущие фактор К в гомозиготном или гетерозиготном состоянии в нормальной цитоплазме, являются потенциальными убийцами; они становятся убийцами фактически, если в их цитоплазме находятся так называемые частички "каппа", которые, в свою очередь, выделяют смертельный яд. В инфузориях типа КК и Кк частички каппа передаются от клетки к клетке в цитоплазме: как только они исчезают из клетки, то не могут возродиться сами по себе. Наследование каппа-частиц вначале считалось великолепным примером цитоплазматической наследственности. Более основательное изучение этого явления, однако, показало, что частицы каппа напоминают бактерии, и их передачу от клетки в клетку в цитоплазме более правильно сопоставлять с передачей паразитических микроорганизмов. То обстоятельство, что частицы каппа могут поддерживаться только в особях с геном К, не противоречит этому доводу. Широко известно из наблюдений, что стойкость или чувствительность к паразиту зависит от генотипа поражаемого хозяина. Имеются доказательства того, что чувствительность к вирусу полиомиелита детерминирована наследственно (см. гл. 16) и что штаммы бактерий могут делаться чувствительными к бактериофагу (см. гл. 25) в результате мутации одного из их генов.
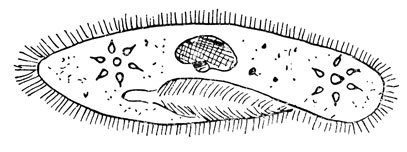
Рис. 92. Микроскопическое животное инфузория-туфелька Paramecium. Бобовидное тело и небольшое, к нему прилегающее тельце представляют собой ядра. Звездообразования - это вакуоли, которые выкачивают лишнюю воду из клетки
У дрозофилы некоторые особи оказываются сверхчувствительными к действию углекислоты и умерщвляются дозами, которые оказывают лишь слабое и временное влияние на нормальных мух. Это свойство передается через цитоплазму; надо опять напомнить, что частицы, о которых идет речь, скорее относятся к вирусам, чем к нормальным составным частям цитоплазмы. Решить, какая из этих двух точек зрения правильна, задача нелегкая. У генетиков нет единого мнения относительно понимания этих и сходных с ними явлений. Даже если их все отнести к истинной цитоплазматической наследственности, общее число этих явлений у животных оказывается чрезвычайно незначительным, и маловероятно, что оно сильно возрастет в итоге будущих открытий. У растений, в которых обратные скрещивания различаются и где отличия не всегда относятся за счет хлоропластов, это явление встречается чаще. Один признак, передающийся у многих растений по наследству через цитоплазму, имеет практическое значение. Это - мужская стерильность, т. е. неспособность образовывать здоровую пыльцу. В тех случаях, например при разведении гибридной кукурузы, когда надо предотвратить возможность самоопыления, внедрение цитоплазматического фактора, создающего мужскую стерильность, экономит труд. Подводя итоги, можно с уверенностью заявить, что в управлении наследственностью монополия генов очень высока, хотя и не абсолютна. Весьма спорным надо считать вопрос о том, велика ли роль цитоплазматической наследственности в определении расхождений сильно отличных друг от друга организмов, принадлежащих к различным видам и родам. К сожалению, этот вопрос не может быть разрешен экспериментально, так как межвидовые и межродовые гибриды обычно бесплодны, а если же и дают потомство, то оно оказывается стерильным (подобно мулу). Мы не будем принимать во внимание цитоплазматическую наследственность в следующей и последней главе - кратком обзоре факторов, играющих роль в процессе эволюции.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'