29. Эволюция глазами генетика
Эволюция - исторический факт. Истинность ее исторически обоснована самым совершенным образом. Доказательства эволюции относятся к различным разделам биологии, например: к изучению ископаемых организмов, данным сравнительной анатомии, к фактам географического распределения животных и растений. Генетика не ставит перед собой задачи обосновать существование эволюции как факт, но стремится объяснить, каким путем шла эволюция. Картина эволюции в генетической трактовке описана во многих, недавно вышедших в свет книгах, некоторые из них написаны в форме, доступной для неспециалиста-биолога; мы можем дать здесь лишь очень краткий очерк вопроса.
Дарвин понимал эволюцию как итог действия отбора на вариации, встречающиеся в природе. Эти вариации возникают в результате случайных процессов и могут быть очень разнообразными; естественный отбор представляет собой силу, направляющую эти вариации по каналу эволюционного прогресса. Принципы естественного отбора уже были рассмотрены в главе 13. Мы сформулируем их здесь следующим образом. Из естественно встречающихся вариаций в любой популяции лучше всего приспособившиеся оставляют более многочисленное потомство, чем остальные. Если свойства, по которым родители оказываются более приспособленными, наследственны, они обнаружатся у потомков, и эти последние опять проявят повышенную приспособленность, что выразится в увеличении среднего числа потомков. В итоге лучше приспособленные особи займут место менее приспособленных, и вся популяция аккуратно приспособится к "нише", создаваемой ее специальным окружением. Эти идеи, впервые выдвинутые в 1859 г, Дарвином в книге "Происхождение видов", были встречены с восторгом и стали руководящими в биологии на многие десятилетия.
Критические высказывания в дальнейшем обращали внимание в основном на проблему природной изменчивости. По Дарвину, вариации, встречающиеся в природе, поставляют материал для селекции. Был задан вопрос о пригодности этого природного материала для осуществления эволюции. Достаточно ли велико число вариаций, возникающих в каждом поколении? Принадлежат ли они к нужному типу? Наконец, возможно, самым главным вопросом надо считать вопрос о передаче признаков вариаций потомству. Если вариации не наследственны, то они совершенно безразличны для процесса отбора, так как, что было показано в главе 21, селекция фенотипических признаков без генетической основы не дает никаких результатов. Возникли еще дополнительные вопросы. Однако, прежде чем о них сказать, попытаемся ответить на вопросы, относящиеся к генетическим вариациям в природных условиях; от решения этих вопросов зависит, признавать или отрицать дарвиновскую теорию эволюции.
Первичным источником наследственной изменчивости надо считать мутации. Как было показано в главе 27, мутация - это процесс, закономерно протекающий в природе. Процесс этот редкий; данный ген может мутировать один раз на миллион клеток или еще реже, скажем, один раз на сто миллионов клеток. Надо, однако, вспомнить, что бактерия, делящаяся на две, каждые два часа даст потомство численностью в сто миллионов клеток в срок меньше трех дней, а мужские половые органы большинства высших организмов дают миллионы сперматозоидов или зерен пыльцы. Кроме того, надо учесть, что эволюция идет на протяжении многих миллионов лет. Таким образом, есть очень много возможностей проявиться любой мутации.
Все же, если сопоставлять успехи эволюции с масштабом времени, выявляется, что одного мутационного процесса едва ли достаточно для образования необходимых наследственных вариаций, если оставить в стороне такие быстро размножающиеся организмы, как бактерии. У более высокоорганизованных форм следующие два фактора строго ограничивают число мутантных особей, возникающих в пределах того или иного отрезка времени: более длинные интервалы между поколениями и более ограниченное число потомков. Эти виды все же должны рассчитывать на мутации как на свой исходный сырой материал: они, однако, используют его, подобно архитектору, который берет строительные блоки разного размера, формы и окраски, чтобы создавать различные сооружения. У этих Видов главным источником генетической изменчивости является рекомбинация, подчиняющаяся второму закону Менделя (см. гл. 19), или для сцепленных генов - процессу кроссинговера (см. гл. 20). Если животновод сталкивается с трудностью или невозможностью разделять или комбинировать два тесно связанных гена (см. гл. 20), то эволюция располагает таким огромным запасом времени, что даже тесное сцепление не создаст серьезных затруднений для процесса перекомбинирования.
Объем генетически обусловленной изменчивости в итоге процесса перекомбинирования колоссален. Рассмотрим четыре пары генов, с которыми работал Мендель: 1 - гладкие семена по сравнению с морщинистыми; 2 - желтые семена по сравнению с зелеными; 3 - высокие растения по сравнению с низкими; 4 - красные цветки по сравнению с белыми. Мы уже видели (см. гл. 19), что рекомбинация первых двух пар генов дает четыре фенотипических класса: гладкие желтые, гладкие зеленые, морщинистые желтые, морщинистые зеленые. В пределах каждого рекомбинационного класса с третьей парой генов подразделение даст растения высокого и низкого роста, так что в целом теперь мы будем иметь 2×2×2=8 различных классов. Рекомбинация с четвертой парой генов приведет к подразделению каждого класса еще на два, в итоге перекомбинирование по всем четырем парам генов даст 2×2×2=16 классов.
Можно идти таким путем и дальше, причем каждая пара аллелей будет удваивать число фенотипических классов. У большинства видов имеются тысячи или десятки тысяч генов, и значительное их число существует в виде двух и больше аллелей. Сделаем допущение, которое надо считать значительно преуменьшенным, что у человека имеется только 100 генов, представленных в виде различных аллелей, и для каждого из этих генов имеется только по два альтернативных аллеля, из которых один рецессивный. Тогда число возможных фенотипов, могущих возникнуть в результате перекомбинирования между этими 100 парами аллелей, будет равно 2×2×2... и так сто раз. Если довести вычисление до конца, получится число, состоящее из единицы с 30 нулями; если это число выразить словами, то придется сказать миллион миллион миллион миллион миллионов. Число генотипов гораздо более многочисленное: каждая пара аллелей (A, a) дает три класса генотипов - AA, Aa и aa, которые при неполном доминировании различаются фенотипически (см. рис. 14).
Рекомбинирование 100 пар аллелей дает 3×3×3 ... и так сто раз, и это выражается числом из единицы с 50 нулями. Так как на Земле живет несколько меньше 3000 млн. человек, очевидно, что практически два человека не имеют никаких шансов быть генетически одинаковыми, за исключением идентичных близнецов. Тот факт, что во всех группах организмов, начиная с бактерий и вирусов, действует половой механизм, позволяющий генам рекомбинироваться, указывает на огромное значение рекомбинирования для процесса эволюции (см. гл. 25). Пол, как и многие другие этапы завоевания Земли жизнью, должен считаться одновременно итогом и причиной эволюции.
Твердо убедившись в том, что в природе широко представлена наследственная изменчивость, мы можем задать себе вопрос, пригодны ли эти вариации в качестве материала для эволюции?
Некоторые ученые высказывали сомнения по этому поводу. Наиболее важные из этих сомнений таковы: 1) мутации являются вредными и не в состоянии объяснить прогрессивную эволюцию; 2) мутации дают резкие отклонения от нормы, тогда как, по Дарвину, эволюция протекает постепенно; 3) мутации захватывают только внешние признаки, вроде окраски глаз; виды же отличаются друг от друга по основным признакам, как анатомическим, так и физиологическим. Эти положения вытекали из ранних генетических открытий, сделанных в начале этого столетия, и в настоящее время устарели. Разберемся в них по порядку.
1. Мы уже познакомились в главе 27 с тем, что большинство мутаций действительно оказываются вредными, если они возникают у вида, который хорошо приспособился к своему образу жизни путем ранее возникших мутаций, рекомбинаций и отбора. Этот факт, вместо того чтобы быть доводом против естественного отбора, скорее является доказательством положения, что в прошлом отбор был эффективен. На протяжении же долгих эпох эволюционного развития условия сами собой меняются относительно каждого вида, и в результате мутаций, которые в прошлом казались бы вредными, могут стать полезными; на самом деле они могут спасти вид от исчезновения. В этом заключается огромное преимущество диплоидии (см. гл. 3), позволяющей потенциально полезным рецессивным генам сохраняться в гетерозиготном состоянии до момента, когда они смогут проявить свою полезность.
Дарвин объяснял преобладание среди насекомых, живущих на островах или на побережьях с частыми ветрами, видов с рудиментарными крыльями, так как при данных условиях существования крылья не нужны и отбираться должны бескрылые мутанты. Французские биологи поставили опыт для проверки этой гипотезы. У дрозофилы рецессивный ген вестиджал превращает крылья в крошечные рудименты. Мухи с мутацией гена вестиджал не могут летать и в нормальной обстановке, где живет дикая дрозофила, поэтому бескрылость является важным недостатком. В местности с сильными ветрами выпустили партии длиннокрылых мух и мух вестиджал. Мух с длинными крыльями ветер относил в сторону с большей легкостью, чем мух вестиджал, и в популяции быстро увеличивался процент короткокрылых мух вестиджал. Если бы в подобных условиях пришлось бы жить дикой популяции дрозофилы, исходно вредная мутация вестиджал могла бы приобрести селекционное значение.
2. Мнение о том, что большинство мутаций проявляется в виде резких изменений, сложилось в генетике исторически. Один из трех ученых, оценивших по-новому работы Менделя в 1900 г. (см. гл. 2), предложил термин "мутация", назвав так резкие наследственные изменения, которые он наблюдал у растения энотера.
На самом деле большинство из них были не мутациями, а рекомбинациями необычного типа. В дальнейшем, в первые годы этого столетия генетики выбирали для установления основных закономерностей наследственности резко выраженные мутационные вариации. Этот выбор подсказывался здравым смыслом; если бы Мендель решил взвешивать каждую горошину, чтобы проследить наследственность незначительных различий в весе, он никогда не открыл бы своих законов. На самом же деле мутации, слабо выраженные, очень распространены; их может обнаружить всякий, пожелавший это сделать, но поисковая работа тяжела и скучна. Совершенно очевидно, что обнаруживают гораздо чаще резко выраженные мутации, чем мутации мелкие; однако нет никаких сомнений в том, что встречаемость последних значительно выше, чем первых.
Особенно существенны для эволюции два типа мелких мутаций. К первому относятся мутации, затрагивающие количественные признаки, вроде размеров тела или плодовитости; как было показано в главе 21, эти признаки управляются суммированным действием многих генов, причем каждый из них проявляет лишь незначительное влияние. Эти гены сами по себе не оказывают заметного действия, но изменяют проявление других, более резких мутаций. Гены-модификаторы играют огромную роль в эволюции, так как на них лежит окончательная отделка - подгонка, полировка и другие операции, подобные тем, которые осуществляют при сборке машины. Два примера могут это проиллюстрировать.
Наиболее обычный тип пегости у мышей зависит от рецессивного гена. Все мыши, гомозиготные по этому гену, несут пегую окраску. Однако степень пегости определяется генами-модификаторами и варьирует от одноцветной окраски до почти белого цвета. У мышей можно повышать или понижать количество белых пятен, разводя мышей с наибольшей пятнистостью или с наименее выраженной пятнистостью, т. е. беря на племя мышей с наибольшим числом генов подходящего рода. В природных условиях естественный отбор часто приводит к более тонким приспособлениям видов к окружающей среде накоплением генов-модификаторов.
Когда обнаруживается впервые новый мутантный признак, он обычно очень резко снижает жизнеспособность его носителя. Богатая практика генетиков показывает, что такие мутировавшие линии могут восстанавливать вполне свою жизнеспособность на протяжении нескольких поколений, при этом отбираются для разведения самые сильные особи. Этот метод разведения приводит к накоплению генов-модификаторов, которые создают "буфер" против неблагоприятного проявления резкой мутации. В процессе эволюции, видимо, процесс буферирования генов играл очень важную роль.
3. Неверное представление о том, что мутации поражают только внешние признаки, несущественные для эволюции, основывается на том, что основатели генетики исследовали законы наследственности на легко наблюдаемых признаках, вроде окраски цветков у гороха и формы крыльев у дрозофилы. Прочитав предшествующие главы, читатель поймет, что мутирование захватывает все признаки, включая самые существенные.
В результате мутирования могут выпадать жизненно необходимые ферменты (см. гл. 23). Наоборот, линии микробов, потерявшие такие ферменты, могут их приобретать вновь путем обратной мутации. Мутации, меняющие особенности цветения растения (см. гл. 16, рис. 16), лежат в основе приспособления к коротким дням в тропиках или длинным дням в умеренной зоне.
У человека иногда встречаются мутации, выражающиеся в появлении перепонок между пальцами на ногах; такие мутации, видимо, играли большую роль в эволюции водных животных. Нет сомнений в том, что мутации, поражающие жизненно важные структуры или функции, должны подчас быть летальными (см. гл. 8), а потому их нельзя обнаружить без специального эмбриологического исследования. Мутации, подавляющие образование позвоночника, известны у мышей, но эмбрионы, гомозиготные по мутантному гену, умирают на самой ранней стадии.
Не приходится сомневаться в том, что мутации, встречаемые в природе и сопровождаемые в большинстве случаев генетической рекомбинацией, представляют собой адекватный и вполне достаточный источник наследственного варьирования, необходимого для осуществления эволюционного процесса. До вторичного открытия менделевских законов этого, однако, не понимали, главным образом потому, что приписывали наследственные отличия отличиям по крови, и скрещивание представляли как результат кровосмешения. Эти термины существуют до сих пор, хотя и потеряли всякий смысл. Такие представления о наследственности приводили к серьезным затруднениям в понимании эволюции с точки зрения Дарвина. Если два окрашенных раствора смешать (красные и черные чернила), их различающие свойства в смеси исчезают. Если бы наследственные признаки находились в крови, они разбавлялись и исчезали бы в конце концов при скрещивании.
Достаточный запас генетической изменчивости может только поддерживаться, если допустить, что новые наследственные отличия будут возникать с высокой частотой. Опыты Менделя одним ударом устранили это затруднение дарвинизма, показав, что наследственные признаки базируются на генах, которые сохраняются в процессе скрещивания и могут проявляться в неизменном виде через очень много поколений.
Дарвин, хотя и был современником Менделя, к несчастью, ничего не знал о его опытах. Он вынужден был вследствие этого не столь определенно высказывать свое представление об эволюции, допуская, что случайное появление наследственных отличий является не единственным источником наследственной изменчивости. В поисках дополнительных источников он обратился к теории Ламарка - знаменитого французского биолога. Ламарка поражала способность организмов приспосабливаться к требованиям их окружения и образа жизни. Кузнец развивает мощные мышцы рук путем упражнений; молодые деревья в густом древостое вырастают высокими в поисках света; мех у пушных зверей зимой более густой, чем летом.
Ламарк допускал, что такие индивидуально приобретенные приспособительные признаки наследуются; так, например, он считал, что у животных, укрепивших мышцы ног под влиянием бега, появится потомство, у которого ножные мышцы с самого начала будут лучше развиты, чем у новорожденного потомства от менее активных (в отношении бега) родителей. Эта теория отвергнута сейчас большинством генетиков.
Попытка экспериментально подтвердить наследование приобретенных признаков никогда не имела успеха. В самом деле, было бы трудно понять, каким образом более сильно развитые мышцы ног могут повлиять на зародышевые клетки, с тем чтобы возникли мутации, обеспечивающие отличное развитие тех же самых мышц у потомков. В то же время невозможно сказать с абсолютной уверенностью, что подобный тип эксперимента никогда не приведет к положительному результату.
Если немногим оставшимся последователям ламаркизма когда-либо удастся получить доказательство наследования приобретенных признаков, это открытие будет обязано как-то приладиться к нашим представлениям в области генетики и эволюции. Однако это едва ли сильно изменит наше современное понимание эволюции. Применение математических методов к изучению эволюции показало, что существующие проявления мутирования, рекомбинации и естественного отбора объясняют вполне удовлетворительно темпы и пути эволюции. Любые дополнительные причины возникновения наследственной изменчивости в лучшем случае могут играть ничтожную роль в поставке материала для грандиозного прогресса жизненных форм от простейших "живых молекул" до ныне наблюдаемого богатства и разнообразия живых организмов.
Имеется еще одно представление, часто привлекаемое к пониманию эволюции. Многие палеонтологи, изучающие ископаемые остатки, считают: как только группа организмов начинает двигаться по новому эволюционному пути, особая направляющая сила начинает следить за тем, чтобы этот путь сохранялся на протяжении многих эволюционных эпох. Легко понять, как возникло это представление.
На рис. 93 показана эволюция передней конечности лошади на протяжении 50 млн. лет. В течение этого времени происходил непрерывный и неизменный процесс перестройки ног этих животных, все лучше и лучше приспособленных к быстрому бегу по безлесным пространствам прерий. В других случаях такая же неумолимая сила, казалось, направляла группы организмов по выбранному ими пути на встречу исчезновению.
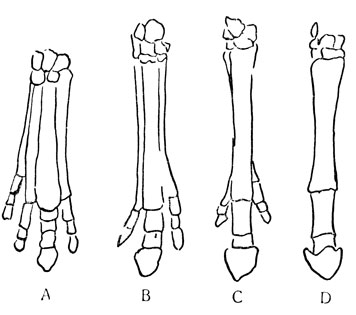
Рис. 93. Эволюция передней конечности лошади: А - более 50 млн. лет назад; В - более 30 млн. лет назад; С - менее 20 млн. лет назад; D - от 1 млн. лет назад до наших дней
Доисторические животные некоторых линий становились все крупнее и крупнее, у них развивались все более и более причудливые рога вплоть до момента, когда линия резко обрывалась. Для негенетиков трудно понять, как могут возникать такие четкие направления на основе отбора, работающего со случайными мутациями и рекомбинациями. Однако мы должны помнить, что хотя мутация и рекомбинация - события случайные, отбор таковым не является. Он направляется двумя условиями: средой, к которой должен приспосабливаться организм, и строением самого организма.
Различные организмы по-разному приспосабливаются к окружающей среде. На одном и том же участке берега можно наблюдать болотных птиц, которые шагают на длинных тонких ногах с широко расставленными пальцами, и уток, ковыляющих по берегу на коротких кривых лапах с перепонками. Этот тип специализации на голенастых и плавающих можно проследить далеко в глубь эволюционной истории птиц. Достаточно было группе птиц с умеренно длинными ногами начать добывать пищу в воде у берегов, любая мутация, способствовавшая утоньшению и удлинению костей ног, давала им преимущества и распространялась по всей группе в результате естественного отбора.
С другой стороны, мутации, укорачивающие кости ног или вызывающие перепонки между пальцами на ногах, оказывались неблагоприятными и скоро вновь исчезали. Чем ближе организм к совершенству по приспособленности, чем меньше число изменений, которые еще могут давать преимущества, тем уже и прямолинейнее становится путь его эволюции. Путешественник может прибыть в далекий город разными дорогами; лишь только он выбрал главную дорогу, возможности пользоваться боковыми дорогами становятся ограниченными, на последних перекрестках его выбор становится несвободным. Эти соображения, наряду с другими, о которых здесь говорить нет возможности, делают излишним для объяснения эволюционных путей обращение к мистической направляющей силе.
Натуралисты, включая и самого Дарвина, отметили еще одну трудность в понимании эволюции путем естественного отбора. Это - наличие у многих видов признаков, лишенных очевидного приспособительного значения, так сказать, "украшательства" эволюции (вроде различных рисунков на крыльях бабочек или различных групп крови у человека). В каждом из таких случаев имеется специфическая трудность. Можно упомянуть лишь два ответа на эти вопросы. Прежде всего признаки могут обладать селекционными преимуществами, которые не выявлены еще для случайного наблюдателя. Английские исследователи эволюции обнаружили, что вариации рисунка из полос на раковинах некоторых наземных моллюсков позволяют им отлично приспосабливаться к разнообразию фона - лесу, канавам, лужайкам, окраска оказывается защитной от нападения дроздов. Большинство генов проявляет плейотропный эффект (см. гл. 23). Бесполезный или даже вредный признак, возможно, управляется геном, обладающим менее заметным плейотропным действием селекционного значения. Разительным примером может служить сохранение гена серповидноклеточной анемии (см. гл. 23) у человека. Новейшие исследования дают основания считать, что группы крови у человека также обладают плейотропным действием, управляя чувствительностью или стойкостью к некоторым заболеваниям. Уместно все же добавить, что нет абсолютно никаких оснований подозревать, что гены окраски кожи человека обладают плейотропным влиянием на какие-либо признаки социального значения.
Остается рассмотреть еще одну важную проблему, внимание к которой было привлечено самим Дарвином. В "Происхождении видов" он задает вопрос: "Если виды произошли от других видов путем незначительных переходов, то почему мы всюду не наблюдаем переходные формы?" Отвечая на поставленный вопрос, среди объяснений он упомянул в качестве важного момента, что "сам процесс естественного отбора непрерывно стремится ...к истреблению ...промежуточных звеньев". Для Дарвина, незнакомого с законом расщепления и рекомбинации генов, это объяснение было вполне приемлемым. Нас оно не удовлетворяет. Мы расцениваем переходные формы как итог случайных генных комбинаций, которые столь же легко разрушаются, как и образуются. Дальнейшая эволюция переходных форм даже в лучше приспособленные формы не могла бы осуществляться без механизмов создания первичных благоприятных генных комбинаций. Эти механизмы носят название "изоляционных механизмов". Как пол и диплоидность, они представляют собой одновременно и результат, и причину эволюционного процесса. Они могут быть весьма разнообразными, но все служат одной и той же цели: ограничение или предупреждение скрещивания и генетической рекомбинации между исходной группой и любой подгруппой, в пределах которой распространился среди достаточного числа особей генотип, обладающий особым преимуществом.
Можно указать лишь на некоторые примеры изоляционных механизмов. У растений мутации, изменяющие время цветения, препятствуют перекрестному опылению мутантной формы и линий, из которой она образовалась. У насекомых - фитофагов (растительноядных) - того же можно достигнуть посредством мутаций, меняющих выбор растения хозяина. У животных мутации, действующие на строение половых органов или поведение при спаривании, могут препятствовать успешному спариванию. Где возможны скрещивания, потомство может оказаться стерильным: лошадь и осел дают жизнеспособное потомство, но их гибриды - мулы и лошаки - оказываются стерильными. Среди изолирующих механизмов наиболее распространена географическая изоляция. Как только достаточное число особей в пределах популяции приобретает новые комбинации генов, они получат возможность заселить новую территорию, где исходные формы не могут выжить.
Вероятно, это происходило многократно в ранние периоды эволюции, когда жизнь впервые вышла из океана на сушу, и произошло вновь, когда обитатели суши (млекопитающие) расщепились на группы, в одной из которых оказались ластоногие (тюлени) и киты, вновь вернувшиеся в океан. В подобных случаях генетические различия предшествовали географической изоляции.
Более часто последовательность оказывается обратной: новый географический барьер (гора, ущелье или пролив) разделял популяцию организмов на две группы, между которыми исчезала возможность скрещивания и обмена генами. В каждой из этих групп естественный отбор благоприятствовал таким особым мутациям и комбинациям генов, которые наилучшим образом подходили к данной среде. Так как нет двух абсолютно одинаковых местностей, это неизбежно приводило к возрастающему генетическому расхождению между этими группами, и это, в свою очередь, приводило к отличиям в строении или поведении, что создавало изолированность, даже если впоследствии барьер исчезал. На этой стадии изоляция осуществлялась полностью. Две группы получают название отдельных видов, если они больше не могут скрещиваться друг с другом и давать плодовитое потомство.
С образованием частично или полностью изолированных групп отбор вступил в новую стадию. Вместо того чтобы воздействовать только на особи, он начал влиять на группы, давая одним возможность распространяться, другие группы хирели и в конечном итоге исчезали. Повсюду, где группы организмов соревнуются за пищу, пространство, свет или за что-либо иное, важное для выживания и размножения, межгрупповой отбор выходит на первое место по сравнению с отбором между особями в пределах групп. Это обстоятельство имело очень важные последствия. Невозможно было оценивать индивид исключительно по его способности давать потомство и передавать гены последующему поколению; гораздо более важным является то, что дает индивид для выживания и способности соревноваться всей труппе. Рабочие пчелы стерильны; тем не менее жизнь и выживание пчелиной семьи зависят от их жизнедеятельности, и там, где семьи диких пчел соревнуются за пищу, решающими являются стерильные рабочие особи, а не плодовитые трутни. В семьях термитов и муравьев высокоспециализированные касты рабочих и солдат возникли путем естественного отбора, действовавшего на группы, а не на особи.
В эпоху расселения человечества успех в соревновании за землю зависел в значительной степени и от индивидуальной плодовитости. Достаточно было общине прочно осесть, ее дальнейшее развитие, способность конкурировать и выживание не зависели больше от плодовитости составляющих ее членов; ведущую роль начинали играть люди разумные, запасливые, энергичные и настроенные в духе общественности. В средние века забота о больных, научная работа и воспитание молодежи находились в основном в руках бездетных монахинь и монахов. В наше время стало поговоркой мнение, что государство с наиболее развитой наукой и техникой будет господствовать на нашей планете; однако ученые и педагоги не могут считаться наиболее плодовитыми членами общества. Агрессивность является преимуществом в соревновании между индивидами, но выживание группы подвергается опасности в случае агрессивности отдельных ее членов. Готовность оказывать помощь, симпатия, самопожертвование приносят вред индивиду в его борьбе за выживание, но благодетельны для группы в целом; в межгрупповом отборе преимущество будет у тех групп, где перечисленные качества развиты сильнее всего.
В отборе, действующем на группы, решающей чертой надо считать общий генофонд группы. (Общий запас генов в группе часто называют "генофондом".) Если группа богата генами, определяющими высокий ПИ (см. гл. 18), в ней будет много интеллигентных людей и средний уровень интеллигентности будет высок; если в группе мало таких генов, средняя интеллигентность будет низкой. Если вернутся к нашему представлению о генах как игральных картах, можно представить себе, что генофонд составляет как бы всю сумму карт, находящихся на руках у людей данной группы. Число карт, конечно, зависит от числа индивидов в данной группе, так как, исходя из правил, игры, каждый человек имеет по две карты каждого рода. Однако пути распределения наборов карт не будут одинаковыми в каждом генофонде и, конечно, будут неодинаковыми, если группы окажутся частично или нацело изолированными. Так, в одном фонде тузы могут быть равномерно распределены по четырем наборам, в другом - бубновые тузы могут встречаться в десять раз чаще, чем тузы трефовые, и в.два раза чаще двух других тузов, взятых вместе; в другом генофонде туз червей может отсутствовать совсем. Особи, рождающиеся в отдельных группах, извлекают карты из генофонда, и это ограничивает и регулирует возможные наборы карт, которые им достаются. Если бубновых тузов в генофонде нет, ни один игрок не может его получить. Если бубновые тузы представлены в десятикратном размере по сравнению с тузами трефовыми, шансы вытянуть их в десять раз выше.
В этой картине степень изоляции двух групп характеризуется частотой взаимообмена двух генофондов картами, т. е. частотой переноса карт из одного генофонда в другой, либо в итоге того, что один индивид перекочевал в другую группу, либо потому, что оба родителя данного индивида происходят из двух разных групп.
Чем чаще генофонды обмениваются картами, тем сильнее будут выравниваться отличия между ними. Если обмен происходит часто, оба генофонда будут содержать сходные наборы карт. Если обмен стал столь частым и любой индивид с одинаковой вероятностью может вытянуть свои карты из того и другого фонда, оба фонда сливаются в общий, в котором карты того и другого фонда скомбинировались в равномерно распределенную смесь. На генетическом языке это можно выразить, сказав, что частота генного обмена при помощи иммиграции и перекрестного скрещивания определяет степень изоляции и в итоге - степень расхождения двух групп. Виды в совершенстве изолированы друг от друга. Между ними нет никакого обмена генами, и их генетические отличия многообразны и глубоки.
Человечество представляет собой обширный вид, подразделенный на группы. Все эти группы могут обмениваться генами путем скрещивания. На этом пути, однако, встречаются препятствия из-за существования более или менее эффективных барьеров между заключаемыми браками за пределами группы. Барьерами могут оказаться географическая удаленность, системы каст и классов и религиозные различия. Когда группы небольшого объема сливаются, например в результате улучшения транспортных возможностей или меньших строгостей в отношении классовой или религиозной принадлежности, вредные рецессивы исчезают с глаз долой, и результат с евгенической точки зрения оказывается положительным (см. гл. 13). Самые крупные и фенотипически отличные группировки человека носят название "рас". Поначалу и в значительной степени до сих пор изоляция между расами была географической.
Там, где исчезли географические барьеры, более или менее строгая изоляция часто сохраняется по причине расовых предрассудков. Древняя и строго проводимая изоляция между расами привела к заметному расхождению их генофондов. Некоторые гены, например гены черной или коричневой окраски кожи, представлены одним отдельным аллелем в некоторых генофондах и иным аллелем в других. Генофонд негров содержит только аллели черной окраски кожи, генофонд европейцев содержит только соответственные аллели белой окраски кожи. В результате действия таких генов каждый индивид одной расы отличается от каждого индивида другой расы. Число же таких генов, однако, очень мало. Многие гены представлены одними и теми же аллелями в большинстве или во всех расах и различаются лишь соотношениями, в которых эти аллели смешаны в генофонде. За исключением некоторых племен американских индейцев, все человеческие расы несут три аллеля групп крови А, В и 0 (см. гл. 10); в то время как аллель 0 чаще всего встречается у всех рас, группа В более распространена среди народов Азии, чем Европы. Точно так же чувствующие на вкус фенилтиокарбамат и нечувствующие (см. гл. 9) были обнаружены среди всех обследованных рас; однако чувствующие вкус ФТК среди североамериканских индейцев встречаются в 94% и только в 70% среди североамериканских белых. По этим генам расы перекрываются, и представители разных рас могут оказаться одинаковыми; и в то же время могут оказаться отличными люди, принадлежащие к одной и той же расе. Мало сомнений в том, что расы различаются также по частоте встречаемости генов, управляющих умственными способностями и личными качествами. Ввиду очень скудных неубедительных доказательств этого положения, которыми мы располагаем, трудно решить, в какой мере это действительно проявляется (см. гл. 18). С другой стороны, нет оснований верить, что какая-то одна раса обладает привилегией носить в себе все высшие аллели таких генов, тогда как другая раса несет в основном самые низшие. Как раз наоборот, имеются все основания считать, что этого нет и что у всех рас встречаются и высшие, и низшие аллели.
Со временем улучшенные способы передвижения и распространение цивилизации неизбежно будут уничтожать генетические барьеры между расами, хотя этот процесс тормозится географическими и социальными препятствиями. Трудно предсказать общий генетический исход всей ситуации. В наше время слишком много детей от смешанных браков не могут успешно развиваться по причине неблагоприятных условий среды. В хороших условиях в семьях смешанных браков рождались прелестные и одаренные дети. Пока дети смешанных браков не будут вырастать в условиях, в которых развиваются дети родителей, принадлежащих к одной и той же расе, трудно оценить по достоинству скрещивание между расами. Уверенно можно говорить лишь об одном. Обмен генами между генофондами, которые долгое время были изолированы, вызовет вспышку новой изменчивости. Если к тому времени человек возьмет в свои руки управление ходом своей собственной эволюции, он сможет использовать это разнообразие для получения великолепных результатов.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'