Глава 2. Взаимодействие генов
8. Что означает полигенность признака? Какие примеры могут проиллюстрировать этот факт?
Полигенность признака - это обусловленность признака многими генами. По-видимому, большинство признаков контролируется многими неаллельными генами, которые взаимодействуют между собой. Простейшими случаями неаллельного взаимодействия являются случаи, когда признак контролируется двумя парами аллелей. Рассмотрим некоторые типы такого взаимодействия.
Комплементарное взаимодействие. Для такого взаимодействия характерно то, что в потомстве от скрещивания двух форм появляется признак, которого не было ни у одного из родителей. Родительские геномы как бы дополняют друг друга и формируют новый признак. Этот тип взаимодействия генов широко распространен и имеет большое значение при изучении комбинативной и мутационной изменчивости.
Например, при скрещивании двух белоцветковых форм душистого горошка потомство оказалось пурпурным, а в F2 шло расщепление: 9/16 пурпурных и 7/16 белых, что соответствовало результатам диаллельного скрещивания. Подобное расщепление является следствием того, что синтез пигмента - сложный процесс, имеющий ряд промежуточных ступеней. Каждый этап этого процесса контролируется определенными ферментами, которые в свою очередь находятся под контролем генов:
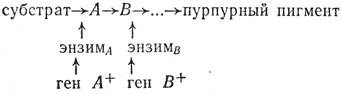
Мутация любого гена блокирует синтез необходимого энзима, и пурпурный пигмент не образуется. Если скрестить две формы, у которых мутация произошла в разных генах, то у гибрида будут синтезироваться все необходимые энзимы, в результате чего образуется пурпурный пигмент:
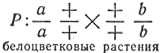

Расщепление по фенотипу 9:7. В F2 формы с блоком одного или двух энзимов белоцветковые.
Такой вид взаимодействия факторов наблюдается и при образовании алкалоидного потомства от скрещивания двух безалкалоидных сортов люпина. Очевидно, у разных сортов безалкалоидного люпина мутация произошла в разных локусах: F1 от скрещивания неаллельных мутантов оказывается алкалоидным, а в F2 наблюдается расщепление 9/16 алкалоидных и 7/16 безалкалоидных. Зная генетическую природу этого явления, его можно предотвратить. Для этого следует проверять сорта люпина, высеваемые в хозяйстве, на совместимость, т. е. на способность образовывать алкалоидные семена при скрещивании.
Новообразование (кооперация). Скрещивание кур породы с розовидным гребнем (виандоты) и кур породы с гороховидным гребнем (брамы) дало в F1 птиц с новой формой гребня - ореховидной, характерной для малайской породы (рис. 3). В F2 наблюдалось такое расщепление: 9/16 - с ореховидным, 3/16 - с розовидным, 3/16 - с гороховидным и 1/16 - с простым (леггорны) гребнями.
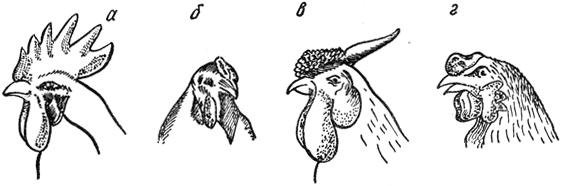
Рис. 3. Формы гребней у петухов: а - простой; б - гороховидный; в - розовидный; г - ореховидный
Итак, в F1 и F2 появляются особи с формой гребня, нехарактерной для пород, использованных в данном скрещивании. Это диаллельное расщепление, при котором простой гребень является результатом взаимодействия двух рецессивов, а ореховидный - двух доминантов (по одному от каждого родителя).
Если R - фактор розовидного гребня, а Р - гороховидного, то получаются следующие формы гребня:

Таким образом, каждая пара аллелей R - r и Р - р наследуется независимо друг от друга, но воздействует на один и тот же признак - форму гребня. Конечный фенотипический эффект зависит от того, какие аллели этих пар присутствуют в генотипе.
У дрозофилы рецессивный ген st-scarlet в гомозиготном состоянии определяет ярко-красную окраску глаз, а рецессивный ген bw-brown - коричневую. Скрещивание этих двух форм дает в F1 нормальных красноглазых мух. В F2 наблюдается расщепление: 9/16 - с красными глазами, 3/16 - с коричневыми, 3/16 - с ярко-красными и 1/16 - с белыми глазами. Окраска глаз зависит от взаимодействия двух независимых пар аллелей:

Если ген st блокирует в гомозиготном состоянии синтез коричневого пигмента, a bw - синтез красного и желтого пигментов, то белая окраска глаз является результатом блокирования синтеза всех пигментов. У форм ±± синтез пигмента идет нормально, что приводит к появлению мух дикого типа по данному признаку.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'