Глава 4. Сцепление и кроссинговер. Карты хромосом
28. Какие признаки называются сцепленными? Как они наследуются?
Любой организм обладает многообразием морфологических, физиологических, биохимических и прочих признаков и свойств, причем каждый признак или свойство контролируется одним либо несколькими генами, локализованными в хромосомах. Однако если число генов организма огромно и может измеряться десятками тысяч, то число хромосом сравнительно невелико (табл. 4). В связи с этим в каждой паре хромосом локализованы сотни и тысячи аллелей, образующих группы сцепления. Установлено полное соответствие между числом групп сцепления и числом пар хромосом. У кукурузы, например, число пар хромосом составляет 2n = 20 и 10 групп сцепления, а у дрозофилы 2n = 8 и 4 группы сцепления.
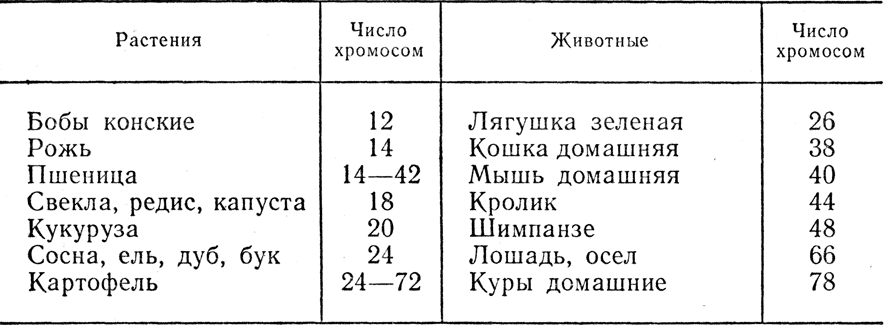
Таблица 4. Диплоидное число хромосом у некоторых растений и животных
Гены, локализованные в одной хромосоме, передаются совместно, и способ их наследования отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом распределении дигибрид A/a B/b образует четыре типа гамет АВ, Аb, аВ, ab в равных количествах, то такой же дигибрид AB/ab при условии полного сцепления факторов А и В (а и b) образует только два типа гамет: АВ и ab. Отклонения от независимого менделевского расщепления как результат сцепления генов впервые установили У. Бэтсон и Р. К. Пеннет (1906 г.). Однако разработка этой проблемы принадлежит школе Т. X. Моргана. Для определения типа наследования двух пар генов (сцепленное или независимое) необходимо провести анализирующее скрещивание:
при полном сцеплении
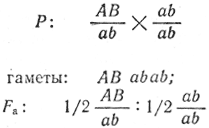
при неполном сцеплении
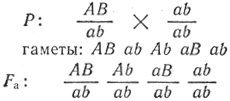
Значит, сцепленные гены могут наследоваться в потомстве как пара аллельных генов. Это справедливо лишь для полного сцепления, когда гены локализуются в хромосоме близко друг к другу. При анализирующем скрещивании в потомстве при неполном сцеплении появляются четыре фенотипа, два из них имеют новое сочетание генов: Ааbb и ааВb. Появление подобных форм свидетельствует о том, что дигибрид с гаметами АВ и ab образует гаметы-рекомбинанты Аb и аВ. Появление таких гамет возможно только в результате обмена участками гомологичных хромосом, т. е. в процессе кроссинговера, и количество их значительно меньше, чем некроссоверных (не рекомбинантных, родительского типа).
Кроссинговер происходит в профазе I мейоза, во время конъюгации гомологичных хромосом. Удвоение хромосом к этому времени завершается, и каждая пара их представлена четырьмя хроматидами. В пахинеме происходит их тесное переплетение, при этом возможны разрыв и объединение хроматид в новые комбинации.
Нужно отметить, что единого мнения относительно механизма обмена гомологичными участками нет: одни исследователи считают, что обмен происходит в момент разрыва хиазм, другие - что хиазмы - это результат уже прошедшего обмена. Однако при отсутствии хиазм кроссинговер обычно не наблюдается.
Так как обмен происходит между участками хроматид, количество кроссоверных гамет не превышает 50%. В редких случаях обмен наблюдается между всеми четырьмя хроматидами, приводя к образованию четырех рекомбинантных хромосом (рис. 9).
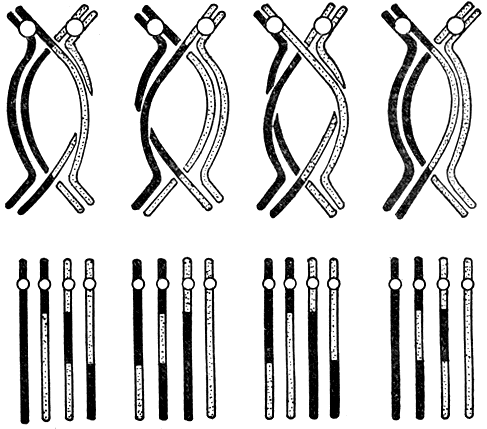
Рис. 9. Схема кроссинговера между различными хроматидами
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'