Глава 10. Решение задач по генетике
190. Как решать задачи по генетике?
Решение задач по генетике способствует усвоению теории и ее практическому применению. Обычно при решении задач проводится генетический или гибридологический анализ. Цели анализа могут быть различными: определение числа генов, контролирующих изучаемый признак, установление характера наследования, определение генотипов и фенотипов потомства или родительских форм и т. д.
Рассмотрим некоторые примеры решения задач на моно-, ди- и полигибридное скрещивание, сцепленное наследование, наследование, сцепленное с полом, неаллельное взаимодействие генов и генетический анализ популяций.
Моногибридное скрещивание
Задача 1. Определить генотипы родительских форм, если при скрещивании желтосемянных растений в F1, F2 и F3 были только желтосемянные. Установлено, что признак - окраска семян гороха контролируется одной парой аллелей, локализованных в паре аутосом.
Решение. Поскольку в течение трех поколений не наблюдалось расщепления, исходные формы гомозиготны.
Задача 2. Определить генотипы родительских форм, если при скрещивании желтосемянного и зеленосемянного растений первое поколение было желтосемянным, а во втором поколении наблюдалось расщепление на 3/4 желто- и 1/4 зеленосемянных.
Решение. F1 было единообразным, значит, родители - гомозиготы. При скрещивании двух различных гомозигот в F1 все потомки должны быть гетерозиготными. Они желтосемянные, значит, этот признак доминирует. Обозначим аллели, определяющие желтую окраску семян, через A, а зеленую - через а. Тогда
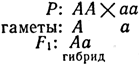
Для получения второго поколения скрещиваем гибриды F1:
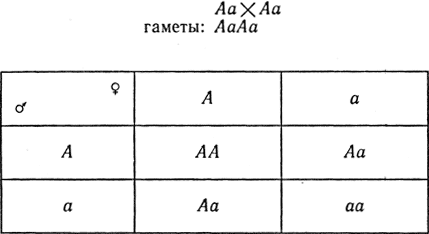
Расщепление по генотипу в F2 1/4 AA + 2/4 Аа + 1/4 аа или по фенотипу 3/4 желтосемянных и 1/4 зеленосемянных.
Задача 3. Какое потомство следует ожидать в анализирующем скрещивании желтосемянных растений их F1?
Решение. Скрещиваем желтосемянное растение F1 с зеленосемянным:
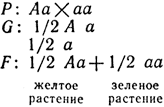
В анализирующем скрещивании получим расщепление 1:1, что является результатом образования двух типов гамет гетерозиготой Аа.
Ди-, полигибридные скрещивания
Если изучается передача двух или более пар признаков, необходимо учитывать, что гены, контролирующие их, могут наследоваться независимо или быть сцепленными.
Независимое наследование
Поскольку каждый признак контролируется одной парой аллелей, локализованных в разных парах хромосом, анализ каждого признака при решении задач должен проводиться отдельно.
Задача 4. Какое потомство следует ожидать от скрещивания гомозиготного желтосемянного растения с зеленосемянным морщинистым, если окраска и поверхность семян гороха наследуются независимо? Желтая окраска доминирует над зеленой, гладкая поверхность семян - над морщинистой.
Решение. Обозначим через A и а аллели желтой и зеленой окраски, а через В и b - аллели гладкой и морщинистой поверхности семян соответственно.
В первом поколении все потомство будет гетерозиготным АаВb с желтыми гладкими семенами, во втором наблюдается расщепление по каждому признаку в отношении 3/4:1/4. Анализируя совместно оба признака, получим 3/4A- + 1/4аа) (3/4B- + 1/4bb):
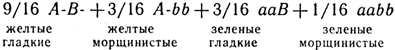
Сцепленное наследование
Если признаки определяются генами, локализованными в одной паре гомологичных хромосом, то они наследуются сцепленно. При полном сцеплении гены всегда наследуются вместе. Благодаря процессу кроссинговера могут возникать кроссоверные гаметы (или гаметы-рекомбинанты), частота которых зависит от расстояния между генами: чем больше расстояние, тем больше вероятность разрывов и обменов и тем больше кроссоверных гамет. Частота кроссоверных и некроссоверных гамет определяет число потомков родительского типа и рекомбинантов, сочетающих признаки матери и отца.
Задача 5. Определить частоту кроссинговера между генами, если при скрещивании серых длиннокрылых мух (дикий тип) с черными короткокрылыми в F1 все мухи были длиннокрылыми, а в анализирующем скрещивании самок F1 с черными короткокрылыми самцами получили расщепление: 722 серых длиннокрылых, 139 серых короткокрылых, 161 черных длиннокрылых и 778 черных короткокрылых. У дрозофилы гены окраски тела сцеплены с генами, определяющими длину крыла. Доминантные аллели серого тела и длинных крыльев обозначим знаком +, черного тела - b, а зачаточные крылья - vg.
Решение. При независимом наследовании в анализирующем скрещивании было бы расщепление 1:1:1:1, т. е. при общем количестве потомков 1800 было бы расщепление 450 серых длиннокрылых, 450 серых короткокрылых, 450 черных длиннокрылых и 450 черных короткокрылых. Но поскольку гены сцеплены, потомство родительского типа значительно превышает число рекомбинантов. Общее число рекомбинантов достигает 300, что составляет 16,7% от общего числа особей в потомстве. Это и есть величина кроссинговера, характеризующая силу сцепления, а значит, и расстояние между генами.
Задача 6. Какое потомство следует ожидать от скрещивания нормальных (серых длиннокрылых) гетерозиготных самок с черными короткокрылыми самцами, если кроссинговер между генами b и vg составляет 16,7%?
Решение. Гетерозиготные самки 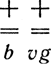
дадут 16,7%
рекомбинантных гамет (151 + vg и 151 b) и 83,3% гамет родительского типа (749 ++, 749 bvg). В задаче были получены другие результаты: 720++, 139 + vg, + 161 b+ и 778 bvg. Разница между теоретически ожидаемыми результатами и результатами, полученными в эксперименте, может быть случайной, т. е. следствием ошибки выборки, неточности данных, заключенных в условии задачи.
Для выяснения причины установленных отклонений полученные данные необходимо обработать по методу χ2.
Составим таблицу:
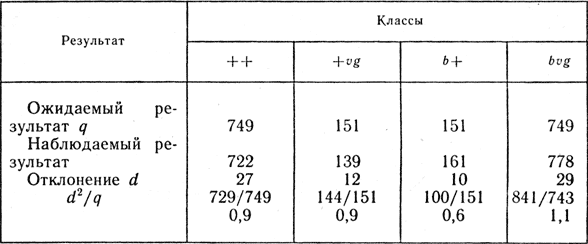
По полученным результатам найдем 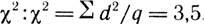
Число степеней свободы при четырех фенотипических классах равно 3. По таблице Фишера найдем 0,2 > Р < 0,5 (см. табл. 1). Значит, отклонение случайное и является результатом ошибки выборки.
Наследование, сцепленное с полом
Задача 7. Допустим, что женский пол гомогаметен (XX), а мужской - гетерогаметен (XY). Например, у дрозофилы окраска глаз - признак, сцепленный с полом, аллель красной окраски обозначен знаком +, аллель белой окраски - w. Какое потомство можно получить от скрещивания белоглазой самки с красноглазым самцом?
Решение. Самка имеет две Х-хромосомы, значит, и два аллеля окраски глаз, самец - одну Х-хромосому, а значит, и один аллель окраски глаз, поскольку Y-хромосома может считаться генетически инертной и не несет аллеля окраски глаз:
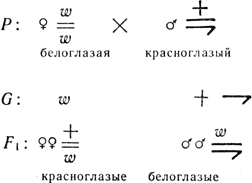
Если один признак сцеплен с полом, а другой не сцеплен, анализ несколько усложняется.
Задача 8. Какое потомство в F1 и F2 можно получить от скрещивания белоглазой длиннокрылой самки с красноглазым короткокрылым самцом, если у дрозофилы Окраска глаз сцеплена с полом, а длина крыльев не сцеплена?
Решение. Обозначим аллель красной окраски глаз через +, белой - w, длинных крыльев - через + и коротких крыльев - через vg. Тогда

Задача 9. Определить порядок генов, расстояние между ними и генотип гетерозиготного родителя, если в анализирующем скрещивании, получено:

Обозначения генов: v+ - красная окраска глаз; v - киноварная; ct+ - нормальные крылья; ct - вырезки на крыльях; cv+ - жилка отсутствует.
Решение. Поскольку полученные результаты явно отличаются от ожидаемых, в случае, если бы гены наследовались независимо, делаем вывод, что наследуемые признаки, а значит, и гены, контролирующие их, по-видимому, сцеплены. Для решения задачи необходимо определить силу сцепления между всеми тремя генами. Выбираем любую пару аллелей, например cv+ ct+, и комплементарную ей cvct и подсчитываем количество особей с избранной комбинацией признаков (73+ , 2+, 2+, 80 = 157). Оставшиеся комбинации этих же аллелей cv + ct и cv ct+ в сумме дают 759 + 140 + 766 + 158 = 1823, что гораздо больше числа предыдущих комбинаций, которые являются рекомбинантными или кроссоверными. Доля кроссоверных особей от общего числа потомков составляет 7,9%. Значит, расстояние между генами будет равно 7,9 единиц рекомбинации, а их расположение на хромосоме окажется таким: cv+ ct или cv ct+.
Аналогичным образом определяется расстояние между следующей парой генов - ct и v. В этом случае сумма ct+v и ctv+ = 73 + 766 + 759 + 80 = 1678, а сумма ct+v+ и ctv = 2 + 140 + 158 + 2 = 202. Значит, расстояние между ct cv равно 10,2% и аллели ct+v расположены на одной хромосоме. Таким же образом определяем расстояние между генами cv и v: [(cv+v + cv v+)] = 73 + 140 + 80 + 158 = 451, что составляет 27,7%. Значит, порядок генов на хромосоме следующий:
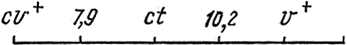
Неаллельное взаимодействие генов
Генетический анализ усложняется, если один признак контролируется не одной, а двумя и более парами аллелей. В таких случаях необходимо установить тип неаллельного взаимодействия и число пар генов (пар аллелей), контролирующих признак.
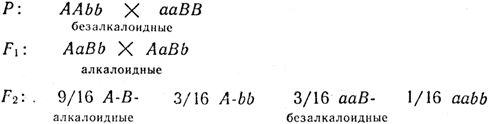
Задача 10. При скрещивании двух безалкалоидных растений люпина в F2 все потомство оказалось алкалоидным, а в F2 наблюдалось расщепление: 9/16 алкалоидных к 7/16 безалкалоидных. Установить генотипы исследуемых форм.
Решение. Родительские безалкалоидные формы генетически отличаются, иначе в F2 они не дали бы расщепления. Они были гомозиготами, так как F1 было единообразным. Наблюдаемое в F2 расщепление свидетельствует о наличии двух пар аллелей (двух генов). Следовательно, формула расщепления у дигибридов 9:3:3:1 в данном случае трансформировалась в 9:7 за счет объединения трех последних классов в один фенотип: 3 + 3 + 1 = 7. Такое расщепление возможно, если для проявления алкалоидности необходимо наличие в генотипе доминантов по обоим генам (АВ), все же остальные генотипы (А-b, ааВ-, ааbb) фенотипически будут безалкалоидными:
В анализирующем скрещивании алкалоидной гетерозиготы с безалкалоидным рецессивом будет непривычное расщепление 3:1:
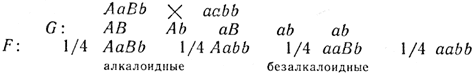
Следовательно, полученное расщепление по фенотипу составит 3/4 безалкалоидных и 1/4 алкалоидных.
Генетический анализ популяций
Решение задач по генетике популяций предполагает выяснение следующих вопросов: какие гены и генотипы представлены в популяции, какова их частота и какой будет популяция в последующих поколениях.
Задача 11. В изоляции оказалась группа особей в соотношении 3АА:6Аа:11аа. Какими будут первое и последующие поколения, если будут сохраняться условия генетического гомеостаза: будут исключены такие факторы, как отбор, мутационный процесс и миграция, и соблюдена равновероятность образования гамет всех типов и панмиксия?
Решение. Определим соотношение аллелей А и а, которое будет соответствовать частоте гамет А и а. Доминантные гомозиготы дадут только гаметы А (3 из 20); гомозиготы аа - только гаметы а (11 из 20); гетерозиготы - 3А и 3а. Значит, частота гамет А окажется равной 3/20 + 3/20 = 6/20 = 0,3; а частота гамет а - 11/20 + 3/20 = 14/20 = 0,7.
Составим решетку Пеннета для определения первого поколения:

F1:9% АА, 42% Аа, 49% аа.
Для определения, каким будет F2, подсчитаем число типов гамет А и а в F1: 9/100 + 21/100 = 30/100 = 0,3 А; 49/100 + 21/100 = 70/100 = 0,7 а.
Составим решетку Пеннета для определения второго поколения:

F2:9% АА, 42% Аа, 49% аа.
Результаты исследований показали, что генетическое равновесие, установленное в первом поколении, сохраняется и в последующих.
Задача 12. Частота людей с группой крови NN в популяции составляет 16%. Найти соотношение групп крови MM, MN и NN и частоту аллелей М и N.
Решение. Учитывая, что частота гомозигот (NN) составляет 16% (0,16), найдем частоту аллеля N в популяции: 
Тогда частота аллеля М составит 1 - 0,4 = 0,6. Определим число гомозигот ММ: 0,62 = 0,36 и число гетерозигот MN: 2 × 0,6 × 0,4 = 0,48 (48%).
Задача 13. Уши кроликов породы баран имеют длину 30 см, других пород - 10 см. Предположим, что различия в длине ушей зависят от двух пар генов с однозначным действием. Генотип баранов - L1L1L2L2, обычных кроликов - l1l1l2l2. Определить длину ушей кроликов в F1 и всех возможных генотипов в F2.
Решение. Длина ушей кроликов относится к количественным признакам, что затрудняет разбивку потомства на четкие фенотипические классы. Генотипы с различным количеством доминантных генов, определяющих длину ушей, будут различными по фенотипу вследствие суммирующего действия указанных генов. Запишем генотип особей F1 и F2:
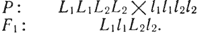
В F2 получим большое разнообразие генотипов с различным количеством доминантных генов (от 0 до 4):

Частота каждого генотипа соответствует коэффициентам разложения бинома Ньютона: 1(0L), 4(1 L), 6(2L), 4(3L), 1(4L) или 1/16, 4/16, 6/16, 4/16, 1/16 соответственно.
Для определения фенотипического выражения каждого генотипа, т. е. длины ушей кроликов различных генотипов, полученных в F2, найдем вклад одного доминантного гена в фенотип: различие между крайними формами (например, у родителей) составляет 20 см при разнице доминантных генов 4. Значит, вклад одного доминантного гена равен 5 см и длина ушей окажется равной 0L - 10 см, 1L - 15, 2L - 20, 3L - 25, AL - 30 см. Формы с двумя доминантными генами (гибриды F1) имеют среднее выражение признака.
Задача 14. В популяции частота дальтонизма (т. е. неспособность различать красный и зеленый цвета) составляет среди мужчин 0,08. Этот дефект обусловлен сцепленным с полом рецессивным аллелем. Каковы ожидаемые частоты трех генотипов у женщин?
Решение. Так как дальтонизм - сцепленный с полом признак, то частота встречаемости этой аномалии зрения у мужчин совпадает с частотой обусловливающего дальтонизм гена q. Следовательно, q = 0,08 и p-1 - 0,08 = 0,92. Тогда ожидаемые частоты различных генотипов среди женщин будут соответственно равны (0,92)2 = 0,846; 2 · 0,92 · 0,08 = 0,147; (0,08)2 = 0,006.
Задача 15. В исходный момент популяция обладает следующей генетической структурой по сцепленному с полом локусу:

Считая скрещивания случайными, рассчитайте равновесные частоты всех генотипов.
Решение. Частоты аллелей в целом по популяции равны:
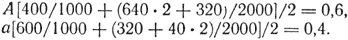
Значит, равновесные частоты генотипов будут следующими:
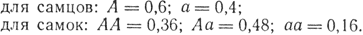
Задача 16. Если частота прямой мутации определенного гена равна 10-5, а частота обратной мутации этого же гена 10-6, то может ли наступить в данной популяции равновесие по этим аллелям и какова будет равновесная частота аллелей?
Решение. Предположим, что частота прямой мутации A1 → A2 = u, а обратная мутация A2-A1 происходит с частотой v. Если исходные частоты аллелей равны р0 и q0, то в следующем поколении частота аллеля А1 будет равна р1 = p0 - up0 + vq0.
Так как доля up0 аллелей А1 превращается в A2, то и доля vq0 аллелей А2 превращается в A1. С учетом этого изменение частоты аллеля А1 за одно поколение обозначим Δр и получим формулу Δр = р1 - р0, в которую подставим значение р1: Δр = (р0 - up0 + vq0) - p0 = vq0 - uр0. Когда Δр = 0, наступает равновесие между прямыми и обратными мутациями. Обозначим равновесные частоты аллелей через р и q и, приняв, что Δр = 0, получим up = vp.
При равновесии число различных аллелей, превращающихся друг в друга, равно. Если р + q= 1 или q = 1 - р, то для равновесной частоты аллеля А1 имеем:
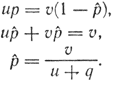
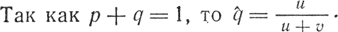
Подставив истинные значения частот прямых и обратных мутаций u = 10-5 и v = 10-6, получим следующие результаты:
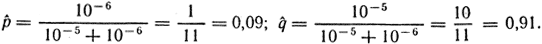
Естественно, что равновесие наступит только при условии отсутствия других возмущающих факторов (миграции, естественного отбора и т. д.).
Задача 17. У кишечной палочки частота мутаций, обусловливающих превращение штамма из не нуждающегося в гистидине в растущий только в присутствии гистидина, и частота обратных мутаций оцениваются следующими величинами: his+ → his- = 2 · 10-6; his- → his+ = 4 · 10-8. Предположив, что никаких иных процессов в популяции не происходит, рассчитать равновесные частоты обоих аллелей.
Решение. Частоты his+ и his- равны соответственно:
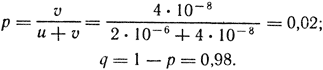
Задача 18. Предположим, что частота мутаций А → а равна 10-6, причем обратные мутации отсутствуют. Какой будет частота аллеля A через 10, 1000, 100000 поколений?
Решение. Частота pt по прошествии t поколений вычисляется по формуле pt = p0(1 - u)t. Если считать, что начальная частота p0 = 1,00, то
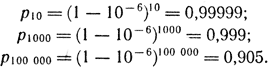
Молекулярная генетика
Задача 19. Искусственно синтезированная информационная РНК (молирибонуклеотид) может служить матрицей при синтезе полипептида. В нескольких экспериментах получены следующие результаты.
Эксперимент I. Поли (А) ведет к образованию полилизина.
Эксперимент II. Поли (АГ) ведет к образованию полипептида, состоящего из чередующихся глутаминовой кислоты и аргинина.
Можно ли по результатам этих экспериментов утверждать, что 1) кодон состоит из 3 оснований; 2) код является перекрывающимся; 3) зная, что код триплетен и неперекрывающийся, какие кодоны кодируют аргинин и глутаминовую кислоту.
Эксперимент III. Поли (ААГ) образует смесь из трех полипептидов: полилизин, полиаргинин, полиглутаминовая кислота.
1. Что можно сказать на основании этого эксперимента об инициации трансляции?
2. Какие уточнения полученных ранее выводов можно извлечь из результатов этого эксперимента?
Эксперимент IV. Поли (АААГ) образует полипептид со следующим чередованием аминокислот: глутаминовая кислота, аргинин, лизин, лизин.
1. Можно ли на основании результатов этого эксперимента говорить о вырожденности генетического кода?
Решение. Результаты эксперимента I могут быть интерпретированы следующим образом:
1. Код моноплетен А - лизин 2. Код биплетен АА - лизин 3. Код триплетен AAA - лизин
На основании эксперимента II можно предположить, что генетический код может быть
1) моноплетным:
А - аминокислота 1 Г - аминокислота 2
2) биплетным и перекрывающимся:
АГ - аминокислота 1 ГА - аминокислота 2 АГ - аминокислота 1
3) триплетным и неперекрывающимся:
АГА - аминокислота 1 ГАГ - аминокислота 2 АГА - аминокислота 1
Следовательно, результаты экспериментов I, II не позволяют определить, является ли генетический код триплетным и неперекрывающимся. Если код триплетен и неперекрывающийся, можно предположить, что аминокислоты - глутаминовая и аргинин кодируются двумя триплетами АГА и ГАГ без уточнения, какой именно триплет соответствует той или иной аминокислоте.
Результаты эксперимента III, в котором на поли (ААГ) синтезируются три разных полипептида (полилизин, полиаргинин и полиглутаминовая кислота), показывают, что инициация синтеза полипептида на такой искусственно синтезированной матрице может начаться с любой точки, т. е. из поли (ААГ) можно получить следующие полинуклеотиды: поли (ААГ), поли (АГА), поли (ГАА), каждый из которых даст начало той или иной полипептидной цепи. Следует подчеркнуть, что такой способ синтеза полипептида может быть только in vitro.
Если бы код был моноплетным, то полученный полипептид включал бы только две аминокислоты, одна из которых была бы представлена в два раза чаще другой: XXYXXYXXY, так как полинуклеотид был бы разбит на кодоны: А|А|Г|А|А|Г|А|А|Г.
Если бы код был биплетным, неперекрывающимся в составе полинуклеотида, были бы выделены следующие биплеты: АА|ГА|АГ|АА|ГА, т. е. был бы получен один полипептид, состоящий из трех аминокислот, чередующихся в такой последовательности: XYZXYZXYZ; если бы код был биплетным, перекрывающимся, мы бы имели следующие рамки считывания: АА ГА АГ|АА, ГА, АГ|, т. е. был бы синтезирован полипептид, состоящий из трех чередующихся аминокислот: XYZXYZXYZ.
Если бы код был триплетным, перекрывающимся на один нуклеотид, мы бы имели следующие рамки считывания: ААГААГААГ (ААГ, ГАА, АГА) был бы синтезирован один полипептид с тремя чередующимися аминокислотами XYZXYZXYZ.
Если бы код был триплетным, перекрывающимся на два нуклеотида ААГААГААГ|(ААГ, АГА, ГАА), то был бы получен один полипептид с тремя аминокислотами. Другими словами, если бы код был моноплетным, биплетным, перекрывающимся или неперекрывающимся, триплетным, перекрывающимся, мы получили бы один, а не три разных полипептида.
Если код триплетен, неперекрывающийся со случайной точкой инициации считывания, он ведет к синтезу трех полипептидов, или,точнее, к трем гомополипептидам.
В эксперименте IV было установлено, что поли (АААГ) служит матрицей для синтеза полипептида, состоящего из глутаминовой кислоты, аргинина, лизина, лизина. С учетом полученной из предыдущих экспериментов информации разбивка на триплеты может быть следующей: AAA ГАА АГА ААГ и т. д. с точкой инициации в любом триплете. Исходя из результатов эксперимента, можно предположить расшифровку кодонов в виде
AAA - глутаминовая кислота ГАА - аргинин АГА - лизин ААГ - лизин.
Однако эксперименты показали, что между кодонами и аминокислотами существует следующая связь:
AAA лизин (эксперимент I) АГА аргинин, глутаминовая кислота (эксперимент II) ААГ
ААГ АГА лизин, аргинин, глутаминовая кислота (эксперимент III) ГАГ
Следовательно, АГА может кодировать аргинин или глутаминовую кислоту. Тогда кодоны ААГ и ГАА будут соответствовать лизину. Итак, лизин кодируется тремя кодонами: AAA, ААГ, ГАА.
В данном случае проявляется вырожденность генетического кода, заключающаяся в том, что одной аминокислоте соответствуют несколько триплетов.
Задача 20. Количество ДНК в ядре клетки человека равно приблизительно 6 · 10-12 г. Известно, что участок двойной спирали ДНК длиной в 0,1 нм имеет массу, равную массе 190 атомов водорода. На основании этих данных определить а) общую длину ДНК в ядре; б) среднюю массу ДНК одной хромосомы и в) длину ДНК одной хромосомы, если допустить, что все хромосомы одинаковы по длине. Сравнить длину ДНК хромосомы и размеры мета-фазных хромосом.
Решение. Масса ДНК длиной в 0,1 нм в 190 раз больше массы атома водорода. Масса одного атома водорода (в граммах) равна 1/6 · 1023. Следовательно, масса 0,1 нм ДНК равна 190/6 · 1023 г. Тогда общая длина ядерной ДНК будет равна:
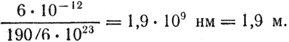
Количество ДНК (в граммах) в одной хромосоме равно:
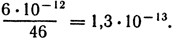
Длина ДНК одной хромосомы
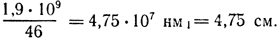
Следовательно, длина хромосомы в интерфазе (длина ДНК) более чем в 40000 раз превышает длину метафазной хромосомы.
Задача 21. Культуру клеток Е. coli равномерно метили 15N, выращивая клетки в течение 14 поколений на среде с 15NH4Cl. Затем эту среду заменяли средой, содержащей 14NH4Cl. При центрифугировании в градиенте плотности ДНК, выделенной из клеток перед пересевом на среду с 14NH4Cl, обнаруживался гомогенный пик ДНК, седиментирующий отдельно от обычной ДНК, которую добавляли в качестве контроля. Этот пик соответствовал ДНК, полностью меченой 15N. После переноса на среду с 14N брали пробы клеток, выделяли в них ДНК и анализировали ее в градиенте плотности. При этом обнаруживали три пика ДНК с различной плотностью. Один соответствовал максимально меченой ДНК, второй - немеченой, а третий являлся промежуточным между первым и вторым. Относительное содержание этих трех ДНК изменялось в зависимости от времени, прошедшего после переноса клеток на среду с 14N:
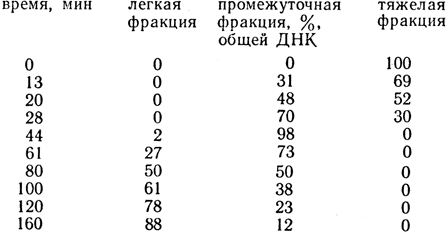
Среднее время генерации клеток было равно 40 мин.
Выполнить анализ полученных результатов.
Решение. В течение первого поколения промежуточный пик увеличивается до 100%, одновременно тяжелый пик уменьшается до 0%. Поскольку это происходит за одно поколение, масса клеток должна удвоиться так же, как и число клеток и количество молекул ДНК. Следовательно, каждая молекула ДНК содержит тяжелый и легкий материал в отношении 1:1. Можно считать, что каждая молекула ДНК содержит одну нить старой ДНК и одну - новой.
В течение следующего поколения количество молекул ДНК вновь удваивается, однако плотность промежуточного пика не уменьшается. Это означает, что в тех молекулах ДНК, в которых после деления содержится тяжелый материал, легкий и тяжелый материал вновь находится в отношении 1:1. Подобный характер распределения материала после деления соответствует тому, что нити ДНК эквивалентны половине молекулы. Если каждая нить в каждом поколении копируется и соединяется со своей копией, то появление легкого пика, образовавшегося во втором поколении, объясняется тем, что легкая нить, синтезированная в первом поколении, соединяется со своей копией, образованной во втором поколении, т. е. ДНК реплицируется по полуконсервативному типу.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'