Генетика и селекция растений

Величайшие открытия сделаны фантастами.
Селекция появилась раньше генетики; прежде она опиралась на отбор, а со временем взяла на вооружение многие методы генетики, биохимии, физиологии и даже математики. Но для этого биологам и селекционерам предстояло открыть гибридизацию у растений.
Гибридизацией биологи называют естественное или искусственное скрещивание различающихся между собой форм. Искусственная гибридизация - один из основных приемов современной селекции. Разработке методов искусственной гибридизации способствовало предшествовавшее развитие представлений о поле у растений.
О существовании пола у растений подозревали, вероятно, еще древние вавилоняне, умевшие искусственно опылять финиковые пальмы. Не сомневались в раздельнополости растений и римские ученые и философы Геродот, Теофраст и Плиний. Но смысла процесса оплодотворения и роли в нем мужских и женских цветков они не понимали, так как наделяли полом и минералы.
Аристотель был убежден, что в растениях женское и мужское начало не отделено друг от друга, и различал растения не по полу, а по плодовитости (плодоносящие растения - женские, не плодоносящие - мужские). Не вполне понял сущность оплодотворения и отец ботаники Теофраст, считавший, например, что плодоносная пальма может быть и мужской, и женской.
Профессору Тюбингенского университета (Германия) Рудольфу Камерариусу (1694) впервые удалось экспериментально доказать наличие мужских и женских органов у растений, использовав в качестве объектов исследований шелковицу, пролесник и шпинат. Английский садовод Томас Ферчайльд был создателем: первого искусственного растительного гибрида, полученного им примерно в 1717 году в результате скрещивания двух видов гвоздики. Гибрид отличался от обеих родительских форм.
К тому времени, когда были начаты широко известные опыты академика Иозефа Готлиба Кёльрейтера (1733-1806), наличие пола у растений было окончательно установлено, доказана возможность и разработана методика искусственной гибридизации. Кёльрейтер получил свой первый гибрид, который назвал бастардом, в 1760 году. В последующие годы он провел многочисленные успешные скрещивания, работая последовательно в Петербурге, Берлине, Лейпциге, Зульце, Кальве и Карлсруэ. К сожалению, Кёльрейтер, несмотря на накопленный богатейший фактический материал, так и не понял значения пыльцевого зерна в процессе оплодотворения, хотя и был осведомлен о роли ветра и насекомых в опылении растений.

Влияние инбридинга на развитие кукурузы (1-8-й год самоопыления)
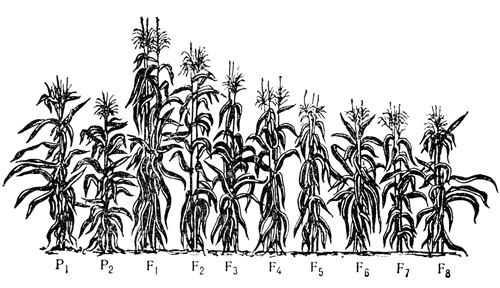
Гетерозис у кукурузы при скрещивании самоопыленных линий (P1 и Р2) и его падение со второго поколения (F2)
Зато он первым обратил внимание на биологический феномен чрезвычайной важности, играющий огромную роль в интенсификации сельского хозяйства нашего времени. Это явление гетерозиса - увеличения мощности, жизнеспособности растения или повышения продуктивности у первого поколения гибридов по сравнению с родительскими формами. Важно и то, что Кёльрейтер указал на значение гетерозиса для растениеводства.
Использование эффекта гетерозиса нашло широкое применение значительно позднее. В двадцатых годах нашего столетия американский генетик Дж. Шелл, используя самоопыление у кукурузы, попробовал избавиться от вредных на его взгляд рецессивных генов. Но растения с каждым новым самоопыленным поколением становились все слабее. Стоило же их скрестить друг с другом, как гибриды (их потомство) приобрели вполне достойный вид. Наиболее экономичным оказалось скрещивание между собой четырех линий (так генетики назвали потомство от многократного самоопыления) по схеме: (AXB)X(CXD), где буквой условно обозначена линия. У таких "двойных межлинейных гибридов" урожай зерна был на 20-30 процентов больше, чем у исходных сортов. Предварительная работа по выведению линии путем ежегодного самоопыления довольно длительная - 8-10 лет, но, если дело поставлено "на поток", вполне себя оправдывает.
Конечно, семеноводам дорогостоящей процедурой представлялась необходимость обрывания вручную метелок с пыльцой у материнских растений. Но и здесь им на помощь пришел генетик. Советский ученый М. И. Хаджинов в тридцатых годах ввел в лучшие линии кукурузы наследственный задаток цитоилазматической мужской стерильности, и необходимость ломать метелки отпала. А позднее генетики позаботились и о том, чтобы сократить период получения самоопыляемых (инбредных) линий. Здесь уже сыграло свою роль явление гаплоидии - обнаружение растений с гаметическим (половинным) набором хромосом, возникающим без участия мужских спермиев. Некоторые гаплоиды естественно удваивали свой набор хромосом. Поскольку они вполне "очищены" от неблагоприятных генов, отпала необходимость в 8-10 годах самоопыления - все свершалось за один год. Затем генетики выделили опылители, которые яйцеклетку не оплодотворяют, но к развитию "без отца" ее все же стимулируют с высокой частотой удачи. Вот только "псевдоотца" пока заменить эффективно не удается ни чужой (иного ботанического рода) пыльцой, ни каким-либо химическим препаратом, ни дорожной пылью (пробовали и так).
У тетраплоидов (а это организмы не с двумя, как у диплоидов, а с четырьмя наборами хромосом) их гаплоиды применяют для гибридизации и отборов на диплоидном уровне с целью быстрого достижения комплексной устойчивости к вредителям, фитофторозу и вирусным болезням, иммунные и сверхчувствительные* реакции к которым контролируются одиночными генами.
* (Сверхчувствительность - тип реакции, при которой инфекция локализуется на определенных участках растения, быстро отмирающих, поэтому широко не распространяется.)
Преимущества селекции тетраплоидов на искусственном диплоидном уровне (например, у картофеля) объясняются тем, что гаметы дигаплоидов (так называют искусственные гаплоиды - диплоиды) содержат аллели в единственном числе (например, А и а), в то время как у тетраплоида могут возникнуть не два, а три типа гамет, содержащих пару аллелей (АА, Аа и аа) и соответственно при их слиянии - три типа гетерозигот (АААа, ААаа и Аааа) против Аа у диплоидов.
Вероятность отбора гомозигот, сочетающих два или более доминантных аллеля устойчивости на диплоидном и тетраплоидном уровнях, выражается следующими отношениями:

Выгода проведения отборов по менделирующпм (подчиняющимся законам Менделя) признакам на диплоидном уровне очевидна. Если предположим, что селекционер ставит задачу отобрать комбинацию, сочетающую в комплексе и гомозиготном состоянии четыре доминантных гена устойчивости к болезням, то такую комбинацию он теоретически может обнаружить у диплоидов, располагая в F2 гибридной популяцией в 256 растений, в то время как у тетраплоидов необходимо не менее 1679 616 растений.
Одиночные гены устойчивости замечательны тем, что их можно относительно легко ввести в культурные растения и защитить последние от рас грибов, особенно досаждающих земледельцам. Вот почему искреннюю радость селекционеров вызвало обнаружение в 1953 году четырех неаллельных доминантных генов устойчивости, обозначенных символами R1, R2, R3 и R4. Каждый ген наследовался независимо и подчинялся (и поныне подчиняется) менделевским законам расщепления в потомстве. Если растение не имеет ни одного доминантного гена, то его может поразить любая раса фитофторы. Чем сложнее раса, тем больший круг сортов она поражает. Так, если раса 1 поражает только растение с фенотипами r, R1, то раса 1.2.4 поражает уже растения r, R1, R2, R4, R1R2, R1R4, R2R4 и R1R2R4, то есть любые растения с генотипами, имеющими в индексе хотя бы одну из цифр, символизирующих расу. Однако чем больше растение имеет R-генов, тем меньшее число рас способно его поразить. Так, фенотип R1 поражают расы 1, 1.2, 1.3, 1.4, 1.2.3, 1.2.4, 1.3.4 и 1.2.3.4, а фенотип R1R2R3R4 - только раса 1.2.3.4.
Сверхчувствительная устойчивость картофеля к фитофторе, обусловленная R-генами, называется вертикальной. В 1956 году этот тип устойчивости растений на примере ржавчины льна был объяснен американским фитопатологом X. Г. Флором на основании гипотезы "ген против гена". Он заподозрил, что у растения и паразита неаллельные (самостоятельно менделирующие), но зависящие друг от друга комплементарные гены. Устойчивое состояние, по Флору, возникает, если взаимодействующие аллели растения-хозяина и патогена (скажем, болезнетворного гриба или вируса) доминантны. Таким образом, если один из взаимодействующих аллелей находится у растения в гомозиготном рецессивном состоянии, оно заболевает. Эти далеко не дружественные отношения растения и паразита, имеющих по одной паре взаимодействующих генов, изображают так:

Плюс - растение восприимчиво, минус - устойчиво.
Вертикальная устойчивость к патогенам, помимо картофеля и льна, обнаружена также у ячменя, овса, пшеницы, кукурузы, томатов, фасоли, яблони и других растений.
Кроме вертикальной устойчивости, фитопатологи выделяют горизонтальную (полигенную) устойчивость к патогенам. Этот тип устойчивости обусловлен целой системой генов, среди которых выделить столь сильные, как R-гены, не удается. Горизонтальную устойчивость патогены преодолевают с трудом, поэтому она выше ценится селекционерами, хотя в процессе селекции труднее достижима главным образом из-за отсутствия хорошего исходного материала. Устойчивость сельскохозяйственных культур, обусловленная R-генами, к сожалению, краткосрочна ввиду легкого возникновения новых рас патогенов.
Устойчивость, обусловленная единичными доминантными генами, широко используется в селекции картофеля на иммунитет к вирусным заболеваниям. Полигенной устойчивостью к вирусам сорта картофеля за редким исключением не отличаются. Во многих странах картофелю как одному из основных продуктов питания отдают предпочтение перед хлебом, потребление которого неуклонно снижается в высокоразвитых странах. Поэтому-то рост числа вирусов, поражающих картофель в ранее "не освоенных" ими районах земного шара, и особенно бурное распространение новых штаммов крайне волнуют селекционеров, так как сводят на нет все их усилия по созданию новых сортов, защищенных доминантными генами. В непрерывной эстафетной гонке патоген - селекционер патогены неизменно побеждали.
И все-таки выход был найден прежде всего благодаря тому, что большинство наиболее вредоносных вирусов передается тлями либо цикадками. Генетиков, естественно, ранее всего заинтересовало, нельзя ли вместо того, чтобы искать генетические источники устойчивости к каждому вирусу (пусть этим продолжают заниматься фитопатологи), взять и включить в селекцию растения, устойчивые к тлям и цикадкам. А такие уже были известны.
На высоте 2-3 тысяч метров над уровнем моря на восточных склонах Анд в Боливии среди чахлых кустарников Качабамбы растет дикий вид картофеля бертхольтии. У этого вида обнаружена очень своеобразная защита не только от тлей и цикадок, но даже от "непобедимого" колорадского жука. Дело в том, что вся ботва растения покрыта волосками, способными при соприкосновении с насекомым выделять клейкое вещество, которое либо намертво приклеивает клещиков, тлю, цикадок и личинок колорадского жука, либо связывает и ограничивает их передвижение.
Солянологи (специалисты, изучающие исключительно семейство пасленовых) давно подозревали, что бертхольтии произошел от естественного скрещивания вида тариензе с каким-то другим. Возник вопрос - нет ли таких особенностей и у них? Особое внимание привлек, конечно, тариензе. Его ареал оказался шире ареала вида бертхольтии. Кроме территории Боливии, он оккупировал еще заросли кустарников и кактусов северо-западной Аргентины.
А в 1971 году профессор Бристольского университета (Англия) Р. В. Гибсон обнаружил подобную особенность еще у одного вида картофеля - полиадениум, произрастающего в Центральной Мексике. Этот картофель превосходил неприхотливостью даже предыдущие два вида, поскольку рос порой на камнях, а то и просто на стенах заброшенных хижин.
Признак клейкости оказался доминантным и контролировался лишь одним геном. А главное, он достаточно легко передавался при скрещивании с другими видами потомству. Селекционеров соблазняло и то, что предварительную селекцию можно было целиком выполнить на диплоидном уровне (это диплоидные виды), вовлекая в работу ценные примитивные культурные виды картофеля или дигаплоиды (диплоиды с генотипом гаметы) столового картофеля. В этом направлении и работают ныне селекционеры Европы и Америки.
К фантастам часто относят селекционеров, берущихся совместить в одном сорте достоинства разных ботанических видов, а то и родов. Возникновения таких форм в природе при естественном переопылении известны и местами не столь уж редки, особенно среди злаковых. Найдены естественные межродовые гибриды пшеницы с дикими злаками - эгилопсом, хайнальдией, пыреем, а также пшеницы с рожью.
Генетиками неопровержимо доказано, что межродовым гибридом диплоидных и тетраплоидных пшениц с видами рода эгилопс является мягкая пшеница - наш главный хлеб. Оказалось, что у мягкой пшеницы (AABBDD) лишь геном А А привнесен пшеницей однозернянкой, а геномы ВВ и DD "подарены" человечеству дикорастущими эгилопсами. И тем не менее межродовые гибриды в природе в целом так редки, что могут рассматриваться как ботанические курьезы.
Созданию межродовых гибридов с участием пшеницы посвятил свою жизнь наш современник академик Н. В. Цицин. Им выделены великолепные промышленные сорта пшенично-пырейных гибридов, опровергшие мнение маловеров о возможности достичь чего-либо путем межродовой гибридизации. На наших глазах происходит становление нового искусственного рода тритикале (AABBBR, где RR от ржи) - первой зерновой культуры, созданной человеком. По мнению специалистов, в недалеком будущем она займет свое место среди ведущих зерновых и кормовых культур.
В 1970 году этой культурой в мире было занято 100 тысяч гектаров, в 1973 году площадь увеличилась в 3,5 раза, в 1975 году уже превышала 500 тысяч. В некоторых сельскохозяйственных районах земного шара по урожайности и питательным достоинствам тритикале превосходит обоих своих родителей - и пшеницу, и рожь. Первый сорт тритикале (Амфидиплоид 1) районирован в 1976 году в СССР. Урожай тритикале в совхозе "Любань" Минской области превысил 82 центнера с гектара. Интересно, что первые шаги в промышленное производство тритикале совершил в 1961 году в Канаде в качестве культуры для производства виски.
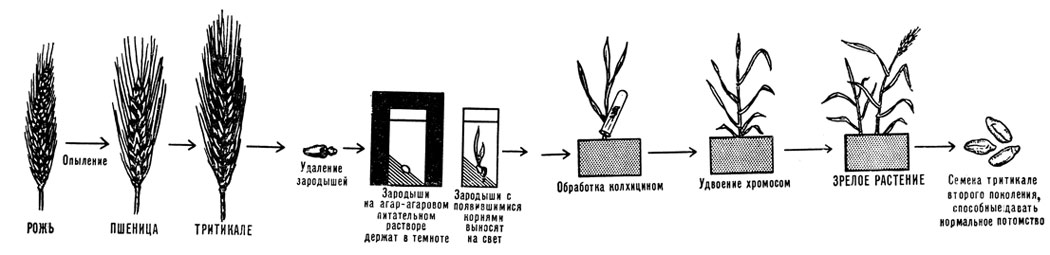
Схема получения искусственного рода тритикале
Пионерами в выведении стабильных пшенично-ржаных гибридов в тридцатых годах были советские исследователи Г. К. Мейстер, Г. А. Левитский, Н. А. Тюмякова и В. Н. Лебедев, которые лишь по не зависящим от них обстоятельствам не сумели довести свою работу до конца. Тем не менее именно их работы явились первыми аккордами грядущего величественного гимна прикладной генетики.
Среди селекционеров-"фантастов" известны имена Н. А. Щибри и И. И. Марченко. Независимо друг от друга они предприняли дерзкую попытку по созданию многолетнего подсолнечника, размножаемого клубнями, для чего скрестили подсолнечник с топинамбуром. Попытка удалась, но, к сожалению, их топинсолнечники существенно уступали по масличности сортам подсолнечника классика мировой селекции В. С. Пустовойта.
Более скромную цель преследовали шведские селекционеры при скрещивании топинамбура с многолетним подсолнечником железистым, дикорастущим в Арканзасе и Миннесоте (США). Они получили гибрид, который назвали Пе-Це-Те (иногда его именуют и Топинэ). Он поразил северян гигантизмом (высота более трех метров) и более высоким, чем у картофеля, урожаем клубней, содержащим 20 процентов углеводов - фруктозы, глюкозы и инулина. Клубни новой культуры хорошо перезимовывают в почве. Урожай зеленой массы - примерно 500 центнеров с гектара - много больше, чем получают от подсолнечника на силос. Зеленую массу можно использовать на корм и в сушеном виде, что облегчает ее транспортировку и хранение.
От скрещивания терна и алычи в результате естественной гибридизации возникла слива. На Кавказе и сейчас можно увидеть дикие гибриды этих видов. Советский ученый В. А. Рыбин повторил эксперимент природы и получил синтетический вид с явными признаками сливы. Среди семенного потомства ученым выделены морозостойкие и быстрорастущие клоны.
Вишня обыкновенная (так ее называют ботаники) - также продукт межвидовой гибридизации в природе. Она появилась в глубокой древности от скрещивания черешни со степной вишней - неказистым кустарничком, не превышающим 1-2 метров. "Опыт" природы удачно воспроизвели английские ученые Е. Олден и Н. Нибон. А вот И. В. Мичурин пошел иным путем, скрестив вишню пенсильванскую - очень морозостойкий в отличие от привычной нам вишни вид - с черемухой и синтезировал новый род, названный по латыни и по-русски церападусом. Лишь много позднее обнаружилось, что церападусы чуть иначе самопроизвольно возникают на Памире.
Блестящие работы по созданию ценных пород межвидовых и межродовых гибридов сливы, вишни, черешни, абрикоса, миндаля выполнены в СССР И. В. Мичуриным и его последователями - И. Н. Рядовым, К. Ф. Костиной, С. Ф. Черненко, А. А. Жаворонковой, X. Е. Еникеевой, А. Н. Веньяминовой, Н. Н. Тихоновой, Н. В. Ковалевой и другими.
Чрезвычайно трудной, однако успешно завершившейся была работа английских селекционеров по гибридизации дикорастущего диплоидного вида ежевики без шипов с тетраплоидной культурной ежевикой, отличавшейся необыкновенно вкусными плодами, но крайне позднеспелой. Вначале исследователям повезло: случайно была найдена ежевика без шипов. Но, несмотря на многочисленные усилия скрестить эти два вида, удалось получить всего лишь четыре сеянца F1 и, увы, все с шипами. Ко всем напастям три из них были триплоидами (с тремя наборами хромосом) и соответственно семян не дали. Но зато последний сеянец, наконец, обрадовал. Он оказался фертильным (плодоносящим) тетраплоидом. Когда дождались плодоношения, посеяли и вырастили новое потомство, было обнаружено, что 37 растений без шипов, а 835 несут шипы. Из первых отобрали одно и, чтобы избежать депрессии, скрестили с культурным, но колючим сортом. В новом потомстве на каждые три растения с шипами пришлось по одному без шипов. Из бесшипных селекционерам приглянулось только одно растение. Оно-то и стало родоначальником знаменитого английского сорта Мертон Торнлесс.
Любимая всеми садовая земляника (ее у нас часто неправильно называют клубникой) произошла от гибридизации земляники чилийской (вообще-то она обитает от Аляски до Патагонии) с земляникой виргинской. И хоть предки ее "американцы", выведена она все же в Европе. В Великобритании в 1819 году Найтом и Кином был искусственно получен межвидовой гибрид, названный в честь одного из авторов Кином.
Очень часто применяют межвидовую гибридизацию в селекции винограда. Пожалуй, даже чаще, чем у картофеля, у которого этот метод селекции является основным (картофель как культуру в Европе от фитофтороза спасло использование в гибридизации дикого вида демиссум). У винограда известны двух-, трех- и даже четырех- и пятивидовые гибриды. Прославленный сорт Дюшес - трехвидовой гибрид, а сорт Вагала - пятивидовой. Предпочитаются, однако, более "простенькие" межвидовые гибриды; у них в меньшей степени проявляются недостатки диких видов. Благодаря включению в селекцию диких видов созданы сорта, устойчивые к поражению филлоксерой и другими вредителями.
Кстати, этот путь селекции винограда открыл И. В. Мичурин. Он первым в мире получил сорта, используя дикие виды. Среди них Буйтур, Арктик и сорт с несколько пугающим названием - Металлический. Другой советский селекционер Н. Н. Тихонов, применив методы И. В. Мичурина, создал межвидовые гибриды промышленного значения для Дальнего Востока.
Широко использовал межвидовую гибридизацию американский селекционер Лютер Бербанк (1849-1926). Пожалуй, одним из самых примечательных его достижений, почему-то почти не упоминаемым его биографами, было создание четырехвидового гибрида карликового съедобного скороспелого каштана, дающего плоды уже на второй год после посева. Вероятно, в селекции древесных пород это единственный пример яркого эффекта селекции по выведению скороспелой формы.
Бессчетное число межвидовых гибридов известно у мандарина. Очень ценными признаны грузинские гибридные сорта помпельмусоидов (мандарин Уншиу Х помпельмус), естественные и искусственные гибридные сорта Вьетнама, Индии, Китая и Японии: танжело (мандарин X грейпфрут), тангоры (мандарин X апельсин), цитрандарины (мандарин Х трифолиата), каламондин (мандарин X кумкват).
Лимон с другими видами цитрусовых скрещивается не так просто, но тем не менее и его удалось принудительно "сочетать браком" с иными видами. Свидетельством тому лимонанжи (лимон X апельсин), лимонаймы (лимон Х лайм), лимондарины (лимон X мандарин). Из всего этого обилия цитрусов жители среднего пояса знают лишь небольшую часть. Культуры очень нежные, не все они могут расти в нашей стране, ведь их родина - тропические влажные леса.
И все же рекордисты среди растений по количеству межвидовых гибридов не цитрусовые, а табак (в основном благодаря американцам, определенно тяготеющим к самому-самому). Только в одном Калифорнийском ботаническом саду проведено 682 межвидовых скрещивания разных видов табака.
Советский селекционер и генетик М. Ф. Терновский широко пользовался в селекции табака межвидовой гибридизацией. Благодаря этому табак как культура был спасен в Краснодарском крае. М. Ф. Терновскому была присуждена высшая из установленных в Советском Союзе премий - Ленинская.
В основном путем межвидовой гибридизации эволюционирует род эвкалиптов. Этим же путем следуют селекционеры. Широко распространяются трехвидовые гибриды. В СССР пользуется успехом как наиболее 90 морозостойкое второе поколение (F2) межвидовых гибридов эвкалипта.
Межвидовые и межродовые сорта-гибриды - равная заслуга прикладной генетики и селекции. И все-таки не эти сорта прославили названные науки в последние десятилетия. Подлинной сенсацией было создание американским генетиком и селекционером лауреатом Нобелевской премии Норманом Борлоугом короткостебельных пшениц сверхинтенсивного типа.
А началось все со случая (вот пример случая, переходящего в необходимость). Борлоуг обнаружил в коллекциях пшениц в США и Международного селекционного центра зерновых культур в Мексике необычайно низкорослую пшеницу, случайно попавшую туда из Индии. Ученый использовал ее в селекции на неполегаемость. Ведь раньше значительные дозы азотных удобрений, вносимых в первую очередь под пшеницу, шли на рост стебля. Вот и получалось - соломы много, а зерна - не очень. А новые карликовые и полукарликовые пшеницы Борлоуга направляли азот главным образом на формирование и налив зерна.
Еще одна карликовая на этот раз японская пшеница была уже известна ученым, но по-настоящему ее "взял в оборот" Борлоуг, обнаруживший у пшеницы целых три гена карликовости. Замечательный селекционер вывел превосходные яровые карликовые и полукарликовые сорта пшеницы, вскоре "оккупировавшие" не только Мексику, но и многие тропические и субтропические страны мира. Урожаи благодаря новым пшеницам Борлоуга и выведенным на их основе (в Индии путем экспериментального мутагенеза) удвоились и даже утроились.
Первые советские полукарликовые сорта пшеницы Полукарликовая 49 и Одесская полукарликовая заполучили гены карликовости от выведенных академиком П. П. Лукьяненко Безостая 1 и мутанта Краснодарский карлик. Вслед за ними на поля пришли созданные в последние годы Донская полукарликовая, Донская безостая, Зирка, Проминь, Мироновская низкорослая и другие высокопродуктивные сорта, имеющие укороченную соломину, устойчивые к бурой ржавчине. Очень хорошие результаты показал в госсортиспытании новый короткостебельный сорт озимой твердой пшеницы.
Гены короткостебельности позволили создать сорта высотой 110-115 сантиметров, устойчивые к полеганию и способные давать 50-60 центнеров зерна с гектара. На значительных площадях возделывается в СССР первый районированный сорт короткостебельной озимой ржи Чулпан, дающий 45-84 центнера зерна с гектара.
Ныне доказано, что рационально возделывать не только низкорослые пшеницы и рожь, но и рис, ячмень, овес, просо и даже сорго и кукурузу на зерно. Вот сколько новых типов полезных растений "подарили" генетики в содружестве с селекционерами сельскохозяйственному производству.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'