Нерасхождение половых хромосом и аберрантные половые типы у дрозофилы
Однако иногда случается так, что в процессе редукционного деления половые хромосомы распределяются между дочерними клетками неправильно. Для примера рассмотрим один такой случай, относящийся к созреванию яиц у нормальной самки дрозофилы.
Неправильность, о которой идет речь, заключается в том, что в первом делении созревания овогония обе Х-хромосомы отходят в редукционное тельце или, напротив, обе остаются в яйце. В результате возникают два новых типа яиц, с которыми мы до сих пор не встречались: в одном из них яйцо вовсе лишено Х-хромосомы (О), в другом - их две (ХХ), т. е. вдвое больше по сравнению с нормой. Нарушение нормального процесса деления клетки, выражающееся в неправильном расхождении хромосом в дочерние клетки и в возникновении гамет иного, чем всегда, хромосомного состава, носит название нерасхождения.
В рассматриваемом случае речь идет о нерасхождении половых хромосом в редукционном делении овогония. Однако необходимо иметь в виду, что такое же нарушение может произойти с любой из аутосом и не только в процессе редукционных делений, т. е. при созревании половых клеток, но и при делении клеток соматических. Несколько подробнее об этом будет сказано ниже.
Проследим теперь типы зародышей (зигот) и взрослых мух, которые возникнут в результате оплодотворения исключительных яиц ХХ + А и 0 + А нормальными сперматозоидами с Х- или У-хромосомой. Возможные при этом четыре типа зигот представлены в табл. 2.
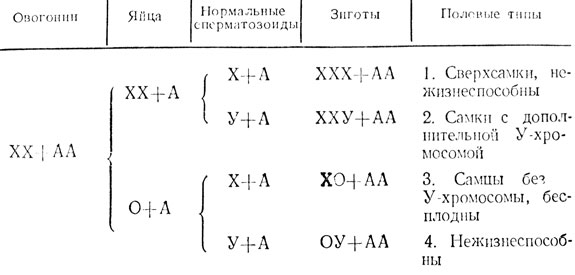
Как видно из этой таблицы, яйца с двумя Х-хромосомами, будучи оплодотворены сперматозоидами с Х-хромосомой, приведут к возникновению самок с тремя Х-хромосомами (ХХХ + АА), а такие же яйца, оплодотворенные сперматозоидами с У-хромосомой, приведут к возникновению самок с двумя Х- и одной У-хромосомами (ХХУ + АА). Первые из них называются сверхсамками, вторые специального названия не имеют и обозначаются как самки с избыточной У-хромосомой.
Следует, впрочем, отметить, что названия "сверхсамки" и "сверхсамцы" (о них будет сказано ниже) с морфологической точки зрения неудачны и даже неправильны, хотя и прочно вошли в терминологический обиход. Они отражают лишь хромосомный состав тех и других и соответствующие сдвиги в значениях полового индекса по сравнению с таковыми нормальных самок и самцов (см. ниже), но не усиление тех особенностей, которые присущи нормальным особям, как то следовало бы из буквального смысла этих терминов. Так, например, сверхсамки ХХХ + АА характеризуются малыми размерами, недоразвитыми яичниками, приводящими к бесплодию, а также нарушениями строения глаз, крыльев, щетинок и другими особенностями.
Самки ХХУ + АА, несмотря на наличие у них дополнительной У-хромосомы, по внешности нормальны и вполне плодовиты. Однако в силу этой причины в их потомстве неизбежно будут возникать исключительные половые типы мух, о чем будет сказано ниже.
Нам остается рассмотреть судьбу зигот и особенности мух, которые возникнут в результате оплодотворения яиц без Х-хромосомы нормальными сперматозоидами с Х- или У-хромосомой.
В тех случаях, когда яйца, лишенные Х-хромосомы, оплодотворены сперматозоидами с Х-хромосомой, из них развиваются нормальные по внешности самцы ХО + АА. Однако отсутствие в клетках этих самцов У-хромосомы приводит к бесплодию; семенники самцов ХО + АА развиты, но содержат лишь незначительное количество неподвижных сперматозоидов.
Что касается яиц, оплодотворенпых сперматозоидами с У-хромосомой, то возникающие из них зиготы ОУ + АА не способны к развитию и погибают.
Таким образом, нерасхождение Х-хромосом в процессе созревания яиц у нормальной самки приводит к возникновению яиц иного, чем всегда, хромосомного состава и, как следствие этого, - к образованию четырех типов исключительных зигот. Из них, за исключением нежизнеспособных зигот ОУ + АА, достигающие взрослого состояния сверхсамки ХХХ + АА и самцы ХО + АА представляют интерес лишь сами по себе, как уклоняющиеся от нормы половые типы. На характер же потомства они не могут оказывать влияния вследствие нежизнеспособности или бесплодия.
Другое дело самки ХХУ + АА с дополнительной У-хромосомой. Однажды возникнув в результате нерасхождения Х-хромосом, они и дальше, во всех последующих поколениях, будут закономерно давать как нормальных самок и самцов, так и мух с нарушенным соотношением половых хромосом и потому ненормальных в половом отношении.
В самом деле, при наличии в овогонии трех половых хромосом - ХХУ - имеются две возможности их расхождения в дочерние клетки в редукционном делении, а именно: Х - ХУ и ХХ - У (табл. 3). В первом случае в редукционное тельце могут отойти две хромосомы, например ХУ, а в яйце останется одна Х-хромосома, или, наоборот, в редукционное тельце отойдет одна Х-хромосома, а в яйце останутся Х- и У-хромосомы. Во втором случае в редукционное тельце может отойти У-хромосома, а в яйце останутся обе Х-хромосомы, или, наоборот, в редукционное тельце могут отойти обе Х-хромосомы, а в яйце останется У-хромосома. Яйца ХХ и ХУ, а также яйца У, если они будут оплодотворены нормальными сперматозоидами с У-хромосомой, снова приведут к возникновению исключительных половых типов и нежизнеспособных зигот УУ +АА.
В табл. 3 приведены возможные типы мух от самки ХХУ + АА и нормального самца. В потомстве этой самки, кроме нормальных самцов и самок (строки 1, 2, 7) и тех жизнеспособных и нежизнеспособных типов, которые были приведены в табл. 2, возникают еще два новых половых типа - ХУУ + АА (строка 4) и УУ + АА (строка 8). Первые представляют собой самцов с двумя У-хромосомами, нормальных по внешности и вполне плодовитых, вторые нежизнеспособны и не достигают взрослого состояния.

Таким образом, между нерасхождением половых хромосом у нормальных самок ХХ + АА и у исключительных самок ХХУ + АА имеется важное отличие; у самок XX + АА оно возникает сравнительно редко, приблизительно в одном овогонии из 2000; у самок же ХХУ + АА нерасхождение половых хромосом будет неизбежно возникать во всех овогониях и в каждом поколении, кроме нормальных мух, давать начало исключительным половым типам, что и было показано на приведенном примере. Для того чтобы подчеркнуть разницу между нерасхождением хромосом у нормальных самок ХХ + АА и у самок ХХУ + АА, первое называют первичным, а второе - вторичным нерасхождением.
Все сказанное относительно нерасхождения половых хромосом в овогенезе в равной степени справедливо в отношении нерасхождения их в сперматогенезе. Возникающие во всех таких случаях исключительные гаметы и половые типы в дальнейшем мы будем называть аберрантными, т. е. уклоняющимися от нормальных в том или ином отношении.
До сих пор, рассматривая механизм возникновения исключительных половых типов на основе первичного и вторичного нерасхождения половых хромосом в овогенезе и сперматогенезе, мы подразумевали, что во всех таких случаях аутосомы распределяются между дочерними клетками нормально и в нормальном числе присутствуют в половых клетках (А) и у всех взрослых аберрантных половых типов (АА). В действительности, однако, правильность расхождения любой из трех аутосом дрозофилы может быть также нарушена, и на этой основе могут возникать исключительные типы мух. В отличие от половых типов, обязанных нерасхождению половых хромосом, их называют аутосомными исключительными типами.
Равным образом нерасхождение половых хромосом и аутосом в сперматогенезе может быть как первичным (сравнительно редким явлением, подобно рассмотренному нерасхождению Х-хромосом в овогенезе нормальных самок), так и вторичным, обусловленным хромосомным составом самой исключительной мухи, т. е. по типу овогенеза у самок ХХУ + АА. Примером могут служить упоминавшиеся выше самцы ХУУ + АА с двумя У-хромосомами.
Сперматогенез у этих самцов будет также протекать с образованием исключительных сперматозоидов ХУ и УУ с последующим возникновением исключительных половых типов, строение которых будет зависеть в том числе и от хромосомного состава оплодотворяемых ими яиц.
Для доказательства нерасхождения аутосом и возникновения на этой основе несбалансированных аутосомных типов нам пришлось бы привлечь другие, более сложные генетические модели и методы, которых здесь мы не касаемся. Поэтому их существование читателю придется принять на веру. Отметим лишь, что в результате воздействия на мух разнообразными внешними агентами и прежде всего рентгеновыми лучами (под влиянием которых разнообразные наследственные изменения и в том числе изменения числа хромосом возникают в большом количестве) и применения остроумных систем скрещиваний у дрозофилы получены и подробно исследованы многочисленные аберрантные типы с нарушенным составом половых хромосом и аутосом. Главнейшие из этих типов приведены в табл. 4.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'