7.2.3. Эволюция белков [1988]
Аминокислотные последовательности белков [51, 81]. Одним из основных достижений биохимии явилось определение аминокислотных последовательностей белков. Гомологичность аминокислотных последовательностей родственных белков стала очевидной вскоре после того, как в конце 1950-х и начале 1960-х гг. были разработаны методы секвенирования. С помощью этих методов была выявлена гомологичность разных, но функционально родственных белков одного и того же вида. По некоторым позициям эти последовательности, как правило, демонстрировали идентичность, а по другим различались. Из результатов изучения ряда вариантов гемоглобина человека в то время было уже известно, что точковые мутации обычно приводят к замещению одной отдельной аминокислоты в полипептидной цепи. В ходе расшифровки генетического кода было показано, что такие замены вызываются замещением одного-единственного основания, происходящим при транскрибировании цепи ДНК. Это открытие стимулировало выяснение эволюционных взаимосвязей между видами путем сравнения числа различий в аминокислотных последовательностях их гомологичных белков. В таких работах строились филогенетические деревья, которые могли сопоставляться с соответствующими схемами, полученными на основе классических палеонтологических и морфологических данных. Методы построения этих деревьев описаны многими авторами [51; 1919; 1921; 1954].
Филогенетическое древо, основанное на результатах сравнения гемоглобиновых генов [1991]. На рис. 7.9 приведено филогенетическое древо для ряда видов позвоночных, включая человека, шимпанзе и гориллу, построенное на основе сравнения аминокислотных последовательностей миоглобина и генов Нbа и Hbp. Временная шкала базируется на палеонтологических данных. Анализ этого древа показывает, что в ходе дивергенции человека и шимпанзе в указанных генах произошла только одна замена основания, а при дивергенции человека и гориллы - три.
![Рис. 7.9. Филогенетическое древо генов мио-глобина, а-цепи и Р-цепи гемоглобина. По ординате отложено приблизительное время расхождения филогенетических линий, установленное на основе палеонтологических данных. Цифры на ребрах - это числа нуклеотидных замен, по которым различаются виды. Цифры в - это числа замен, скорректированные на многоступенчатые мутации. Последовательность миоглобина свиньи пока расшифрована неполностью, поэтому соответствующая оценка числа замен слегка занижена [81]](pic/000015.jpg)
Рис. 7.9. Филогенетическое древо генов миоглобина, α-цепи и β-цепи гемоглобина. По ординате отложено приблизительное время расхождения филогенетических линий, установленное на основе палеонтологических данных. Цифры на ребрах - это числа нуклеотидных замен, по которым различаются виды. Цифры в ▫ - это числа замен, скорректированные на многоступенчатые мутации. Последовательность миоглобина свиньи пока расшифрована неполностью, поэтому соответствующая оценка числа замен слегка занижена [81]
Похожие схемы можно построить и для других белков, а также, объединив соответствующие данные, для совокупности всех известных белков. Они могут охватывать не только позвоночных, но и более широкий круг таксонов, включающий беспозвоночных и даже (при использовании таких универсальных белков, как гистоны или цитохром С) растения, грибы и микроорганизмы.
Темпы эволюции разных белков. Можно провести сравнение числа мутаций, зафиксированных в разных белках при прохождении определенного числа этапов видообразования. Оказывается, что некоторые белки эволюционируют с гораздо большей скоростью, чем другие (табл. 7.4). Гистоны, например, удивительно стабильны, тогда как эволюция фибринопептидов шла очень быстро. Учитывая, что в ходе эволюции фиксируется лишь незначительная часть всех мутаций - приблизительно одна на 3,5 миллиона [1914], - мы можем без колебаний отвергнуть объяснение, связывающее различия в скорости эволюции с различиями в частотах возникновения мутаций; по-видимому, причину правильнее искать в функциях соответствующих белков. Например, функция фибринопептидов не является очень специфичной (они отщепляются в процессе превращения фибриногена в фибрин). Этим, возможно, и объясняется высокая скорость их эволюции. С другой стороны, конформация гистонов, очевидно, подвергается очень сильным ограничениям. Они вступают в тонкое пространственное взаимодействие с ДНК. Весьма вероятно, что эти белки выполняют какую-то критическую функцию, которая нарушается даже при небольших изменениях в их молекулах.
Дупликации генов. Как отмечалось в разд. 4.3, молекула гемоглобина А (НbА) состоит из двух α- и двух β-цепей; HbF вместо β-цепей содержит γ-цепи, a HbA2-5-цепи. Во многих гомологичных сайтах цепей всех четырех типов находятся одинаковые аминокислоты. Наиболее очевидное объяснение данного факта состоит в том, что все эти гены, а также ген цепи миоглобина, произошли от одной предковой последовательности. Для такой функциональной дифференциации требовалась дупликация этих генов. В результате одна из копий продолжала выступать в прежней роли, в то время как другая имела возможность приобрести новую функцию. На рис. 7.10 показаны этапы дуплицирования гемоглобиновых генов, приуроченные к стадиям эволюции и привязанные с известным приближением к временной шкале.
![Таблица 7.4. Скорость фиксации мутационных замен в ходе эволюции [51]](pic/000016.jpg)
Таблица 7.4. Скорость фиксации мутационных замен в ходе эволюции [51]
* (1 ФТМ-1 фиксированная точковая мутация/100 аминокислотных остатков за 100 млн. лет. Фиксированной точковой мутацией называется замена в данном белке одной аминокислоты на другую.)
Дупликации генетического материала - отдельных генов, коротких участков хромосом и генома в целом (полиплоидизация) - имеют ключевое значение для эволюции. В ходе эволюции млекопитающих полиплоидизация, по-видимому, уже не происходила [157]; в то же время небольшие дупликации и нехватки возникали довольно часто.
Эволюция генов, кодирующих белковые домены. До сих пор мы рассматривали только изменения в аминокислотных последовательностях. Известно, однако, что белки имеют специфическую трехмерную структуру, которая обычно создается двумя или более расположенными друг за другом "доменами", т. е. последовательностями с молекулярной массой в 20000, свернутыми таким образом, что количество контактов внутри домена оказывается намного больше, чем между последовательностями разных доменов. При сравнении доменов из разных белков обнаружилось, что конформационное сходство распространено существенно шире, чем ожидалось на основании результатов сопоставления аминокислотных последовательностей. Белковые домены могут иметь очень похожие конформации и при отсутствии сходства в аминокислотных последовательностях. В ходе эволюции фиксация мутаций происходила только в том случае, если соответствующая аминокислотная замена не нарушала конформации белка [1979]. Согласно теоретическим расчетам, всего 200-500 доменов могли послужить основными единицами, из которых составлено громадное число различных белков, имеющихся у живых организмов. Как отмечалось в разд. 2.3.3, гены эукариот состоят из нескольких экзонов (экспрессируемых последовательностей ДНК), разделенных нитронами (неэкспрессируемыми последовательностями). Отдельные экзоны, по-видимому, часто содержат последовательности ДНК, кодирующие такой белковый домен.
Например, можно показать, что ген фактора VIIIC (разд. 2.3.3.7) соответствует трем доменам, обозначаемым буквами А, В и С. Домен А состоит из 330 аминокислот, домен В - из 980, а домен С - из 150. Эти домены располагаются в порядке А1 - А2 - В - A3 - С1 - С2. Домен А обнаруживает неожиданную, но "значительную гомологию с церулоплазмином" - медьсодержащим белком сыворотки, который также имеет три домена А, но не содержит доменов В и С (см. разд. 2.3.7).
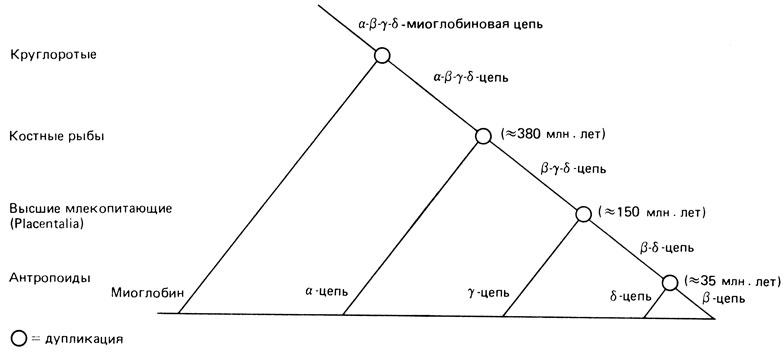
Рис. 7.10. Этапы дуплицирования гемоглобиновых генов и стадии эволюции, на которых происходили дупликации. Дополнительные дупликации привели к появлению гемоглобиновых цепей, обнаруживаемых у ранних эмбрионов. Кроме того, у человека имеется еще по две γ- и α-цепи. Информация о точном времени дупликаций в случае генов этих цепей отсутствует
Полезные или нейтральные мутации? Почему определенные аминокислоты в последовательности за длительный период замещаются другими? Существуют два возможных объяснения: либо эти замены приводят к усовершенствованию функциональных свойств молекулы, дающему ей селективное преимущество, либо мутации нейтрал! ты или даже слабо вредны и фиксируются в результате генетического дрейфа или случайно (разд. 6.4). Последнее объяснение особенно ревностно отстаивает Кимура [1941; 1942].
Во введении к своей монографии [1941] Кимура пишет: "Теория нейтральности утверждает, что большинство эволюционных изменений на молекулярном уровне, выявляемых при сравнительном изучении аминокислотных последовательностей белков и нуклеотидных последовательностей ДНК, обусловлено не дарвиновским отбором, а случайным дрейфом селективно нейтральных или почти нейтральных мутаций. Эта теория не отрицает роли естественного отбора в определении направления адаптивной эволюции, однако она предполагает, что адаптивную природу имеет лишь незначительная часть эволюционных изменений первичной структуры ДНК, тогда как громадное большинство фенотипически "молчащих" замен нуклеотидов не оказывает никакого существенного влияния на выживание и воспроизведение...
В теории нейтральности утверждается также, что большая часть внутривидовой изменчивости на молекулярном уровне, проявляющейся, например, в виде полиморфизма белков, нейтральна, и поэтому большинство полиморфных аллелей, имеющихся у какого-либо вида, поддерживается за счет мутационного процесса и случайной элиминации. ... (Она) трактует полиморфизм белков и ДНК как переходную фазу молекулярной эволюции и отвергает представление, согласно которому большинство таких систем полиморфизма имеет адаптивное значение и в пределах вида поддерживается одной из форм балансирующего отбора".
С другой стороны, Кимура проводит ясное различие между положительным (приспосабливающим) отбором, который, как он полагает, очень редко действует на молекулярном уровне, и отрицательным отбором, благодаря которому происходит элиминация большого числа вредных мутаций. Он даже дает оценку, согласно которой нейтральными (или слабо вредными) могут быть лишь ≈ 10% всех новых мутаций; ≈ 90% новых мутаций относятся к категории безусловно вредных и не имеют шансов зафиксироваться в популяции.
Гипотеза "нейтральности" вызвала жаркие дискуссии среди генетиков-популяционистов. Отчасти они вызваны неправильным пониманием отдельных положений данной гипотезы, что убедительно разъясняется самим Кимурой [1941].
Чтобы понять эту гипотезу, необходимо уяснить два следующих момента.
1. В 1960-е годы была обнаружена громадная генетическая изменчивость на уровне белков и соответственно ДНК. С помощью методов определения аминокислотных последовательностей удалось выявить различия между гомологичными белками разных видов, а также между родственными белками одних и тех же видов. Изучение генетического кода вскрыло новые источники изменчивости, нуждающиеся в дальнейшем исследовании. Огромное количество ДНК, обнаруженное в эукариотической клетке (разд. 2.3.1.1), породило вопрос о функции избыточной ДНК и возможной причине этого феномена. Связаны ли большое количество ДНК и ее значительная изменчивость с естественным отбором, как это предполагалось неодарвиновской теорией эволюции, или же на молекулярном уровне большее значение имеют случайные процессы? Если бы решающим фактором был, как это предполагалось общепринятой синтетической теорией, отбор, то его действие испытывало бы огромное число сайтов ДНК.
2. Кимура разработал математические диффузионные модели, которые позволяют получить ответ на вопрос: "Какова вероятность того, что отдельный мутант, возникший в популяции конечного размера, рано или поздно распространится по всей популяции?" (т. е. оценить вероятность фиксации гена, разд. 6.4.1 и 6.4.2).
В своей классической работе он сформулировал эту проблему и ее решение следующим образом:
"Рассмотрим популяцию численностью в N особей. ... Если мы заглянем достаточно далеко в будущее, то увидим, что вся популяция генов данного определенного локуса происходит от одного-единственного аллеля, имеющегося в нынешнем поколении. Это результат фатального процесса случайного генетического дрейфа*. Если в нынешнем поколении аллель А1 имеет частоту p, вероятность того, что удачливым аллелем, от которого произойдет вся популяция генов, будет А1, а не какой-нибудь другой аллель, равна ... р.
* (Это положение следует из законов, лежащих в основе процесса случайной фиксации; разд. 6.4.)
Теперь, если мутация возникает с частотой μ на ген на поколение, то число новых мутантов по этому локусу в нынешнем поколении составляет 2Nμ*. Кроме того, вероятность, что данный ген рано или поздно зафиксируется в популяции, равна 1/2N. Таким образом, вероятность возникновения в этом поколении какого-либо мутантного гена и его фиксации в данной популяции равна
* (Где N = размер популяций.)
| 2Nμ× | 1 | =μ, |
| 2N |
т. е. независимо от размера популяции частота нейтральной генной замены равна частоте мутаций".
Позднее Кимура несколько скорректировал этот тезис, заметив, что мутант, селективно невыгодный в большой популяции, может быть нейтральным в небольшой и, таким образом, в малых популяциях скорость замен на самом деле будет выше.
В своих модельных расчетах Кимура рассматривал случайное мутирование как процесс, независимый от времени. В широко известной монографии [1941] он пишет, что основываясь на положении о случайной фиксации мутаций, зависящей только от их частоты, можно сделать предсказание о линейности накопления генетических различий с течением времени, существующей независимо от изучаемого вида, продолжительности его поколения и других параметров. Пределы скоростей фиксации замен определялись только ограничениями, налагаемыми функциональными требованиями к генам и их продуктам - белкам: "отрицательный" отбор элиминировал макромолекулы, содержащие замены оснований и аминокислот, не совместимые с нормальным выполнением функции. Различия скоростей эволюции белков (табл. 7.4) могут в полном соответствии с общепринятой теорией объясняться этим отрицательным отбором. Единственное, что она отвергает, - это объяснение существования различных последовательностей, поддерживаемых в ходе эволюции, соответствующими различиями в действии положительного отбора.
Справедливость и значение гипотезы нейтральности для понимания определенных аспектов эволюции можно оценить, выяснив, подтверждают или опровергают имеющиеся реальные данные те следствия, которые вытекают из этой теории. Они касаются эволюционных изменений на уровне аминокислотных последовательностей (т. е. аминокислотных замен в белках) и на уровне нуклеотидных последовательностей (т. е. замен оснований в ДНК).
Как уже отмечалось, одно из важнейших следствий гипотезы нейтральности - это положение о линейной зависимости частоты накопленных замен от времени, т. е. предсказание существования молекулярных часов эволюции. Для проверки концепции эволюционных часов требовались оценки времени разделения соответствующих ветвей филогенетического древа, полученные с использованием независимой шкалы времени, основанной, например, на палеонтологических данных. Участники ведущихся в литературе дискуссий относительно рассматриваемого следствия из гипотезы нейтральности часто оперируют этими данными. Для обоснования концепции эволюционных часов приводят, например, следующий аргумент: некоторые глубоководные виды рыб с незапамятных времен обитают в океане и экологические условия их обитания должны быть очень сходны, если не одинаковы, на протяжении всего времени их существования; тем не менее эволюция белков этих рыб протекала с постоянной скоростью, α- и β-цепи гемоглобина у млекопитающих, отличных у человека, со времени разделения дивергировали в такой же степени, как α- и β-цепи гемоглобина человека и гемоглобина рыб. С другой стороны, более детальные исследования некоторых частей "филогенетических древ" выявили отклонения от линейной зависимости: например, эволюция белков приматов протекала медленнее, чем ожидалось на основании гипотезы молекулярных эволюционных часов.
Аргументы против универсальности гипотезы нейтральности. Дискуссия о гипотезе нейтральности развернулась и в теоретической популяционной генетике. Достаточно сказать, что основываясь на конкретных допущениях, можно прийти к выводам, отличным от тех, что были получены Кимурой (см., например, работу Ивенса [1757]). В идеале теория и вытекающие из нее положения должны проходить экспериментальную проверку. К числу бесспорных относятся следующие тезисы:
1) многие виды мутаций приводят к генетическим дефектам. Однако такие мутации селективно вредны и быстро элиминируются из популяции;
2) другие мутации подвергаются специфическим формам отбора, поддерживающим генетический полиморфизм либо в результате гетерозиса (разд. 6.2.1.3), либо посредством частотно-зависимого отбора (разд. 6.2.1.5);
3) многие аминокислоты, локализованные в тех или иных сайтах белков, в современных условиях не имеют никакого ощутимого селективного преимущества по сравнению с аминокислотами, которые они заместили в ходе эволюции.
Возможно, что какая-то доля мутаций в период фиксации все же обладала слабым селективным преимуществом. Некоторые недавно полученные данные свидетельствуют о следующем:
1) если функция какого-либо генного продукта уже установилась, отбор будет стремиться сохранить функциональные характеристики или даже несколько их улучшить. Поэтому замена одной аминокислоты на другую, имеющую сходные конформационные и биохимические свойства, должна поддерживаться отбором [1509; 1907]. И действительно, существует сильная корреляция между биохимическим сходством аминокислот и вероятностью замещения;
2) если мутант возникает, скажем, в результате дупликации, то ожидается, что отбор будет приспосабливать его к выполнению какой-то новой функции; при этом частота данной аминокислотной замены, по-видимому, будет расти. Это положение основано на данных о гемоглобинах, частота аминокислотных замен в которых после возникновения дупликаций увеличивалась [1927]. Заметим, однако, что некоторые исследователи высказывают сомнения в достоверности палеонтологических данных, на которых базируются филогенетические древа [1941];
3) гипотеза нейтральности утверждает, что многие (а возможно и большинство) существующие в современной популяции человека системы полиморфизма не поддерживаются отбором и представлены нейтральными аллелями, находящимися на пути к фиксации посредством случайного дрейфа.
На первый взгляд кажется, что распределение редких и обычных электрофоретических вариантов противоречит этому постулату (см. разд. 6.1.1; рис. 6.4). Наблюдаемое распределение строго бимодально; существует группа с относительно высокими (промежуточными) генными частотами, которая, вероятно, поддерживается за счет преимущества гетерозигот и частотно-зависимого отбора. Кроме того, имеется группа с низкими частотами, в нее могут входить гены, не обладающие каким-либо селективным преимуществом или вредностью и поддерживаемые посредством дрейфа. Гипотеза нейтральности предсказывает относительно высокую частоту различных редких и обычных (частота гена ≥ 0,9) вариантов и более низкую частоту вариантов, встречающихся с промежуточными частотами; такое распределение, очевидно, отличается от фактического, построенного на основе реальных данных. Однако пока очень трудно установить, что именно обусловило характер полученного распределения - преимущественно генетический дрейф нейтральных аллелей, сочетание различных форм отбора или же и то и другое вместе;
4) эта гипотеза может также обсуждаться на материале, полученном при изучении ДНК. Например, замены оснований, не приводящие к замещениям аминокислот (особенно оснований в третьих позициях кодонов), являются, как было обнаружено, более распространенными, чем замены, вызывающие такие замещения; особенно вариабельными оказались последовательности ДНК нетранскрибируемых районов. Эти положения относятся также и к внутривидовой изменчивости человека, у которого описано множество систем полиморфизма по сайтам рестрикции ДНК (разд. 2.3.3.9 и 6.1.2). Согласно недавно полученной оценке [328], средняя гетерозиготность на кодон по некодирующим последовательностям ДНК генома человека, вероятно, примерно в десять раз выше, чем по кодирующим. Кроме того, скорость замещения оснований в функционально инертных псевдогенах, например в псевдогене Нbа мыши (разд. 4.3), по-видимому, выше, чем в его активных дубликатах [1953]. С другой стороны, сравнение мРНК β-цепей гемоглобинов человека, мыши и кролика не дало каких-либо указаний, свидетельствующих о случайности замен, ожидаемой на основании гипотезы нейтральности; напротив, характер распределения замен оснований был явно неслучайным [1918]. Многие данные, использовавшиеся для обоснования и опровержения гипотезы нейтральности, можно трактовать по-разному. Пока все эти вопросы не нашли однозначного ответа, попытаемся сформулировать некоторые правдоподобные выводы.
"Генетическая достаточность" [1992; 1993]. Предположим, что условия окружающей среды изменяются таким образом, что функциональная адаптация определенного полипептида становится менее эффективной. Тогда, если происходит мутация, лучше удовлетворяющая новым требованиям, ее носители будут иметь селективное преимущество. Эта новая мутация не обязательно усовершенствует данный полипептид до наиболее оптимального состояния; преимущество может оказаться совсем небольшим. Важно помнить также, что любое усовершенствование может быть обусловлено рядом различных мутаций; природа имеет в запасе много вариантов ответа на каждое требование, не всегда оптимальных, но часто вполне адекватных. Отбирается тот мутант, который присутствует в популяции в то время, когда среда предъявила этому полипептиду свои новые требования. Наличие мутантов в свою очередь зависит от частоты возникновения мутаций (и от генетического дрейфа). По некоторым данным, мутации, зафиксировавшиеся в ходе эволюции, чаще являются результатом транзиций оснований, чем трансверсий; транзиций более распространены по-видимому, и среди новых мутаций (разд. 5.1.4) [1985]. Следовательно, элемент случайности проявляется здесь в пределах ограничений, налагаемых на данный белок требованиями, выполнение которых необходимо для его нормального функционирования, и отбором.
Смысл понятия "генетической достаточности" можно пояснить на одном примере, известном специалистам по генетике человека. Когда в местных тропических странах широко распространилась малярия, для популяций стала полезной повышенная устойчивость к этой болезни. Вскоре во всех таких популяциях возникли определенные механизмы генетической адаптации. Однако в разных популяциях конкретные способы адаптации были различными (разд. 6.2.1.6). В Африке происходил отбор HbS и НbС, а в австрало-азиатской популяции - НbЕ; в некоторых других популяциях повысились частоты талассемий и различных недостаточностей глюкозо-6-фосфат-дегидрогеназы. Адаптивные величины этих мутаций отнюдь не одинаковы; НbЕ, например, обеспечивал защиту от малярии "за значительно более низкую цену", чем β-талассемия, поскольку у гомозиготных носителей генов НbЕ патологические симптомы выражены слабее, чем у гомозигот по генам β-талассемий (разд. 6.2.1.7). Тем не менее обе адаптации оказались достаточными, так как эти популяции выжили. Очевидно, что возникновение той или иной адаптации зависело от типа мутации, которая имелась в наличии и поэтому могла быть поддержана отбором.
Концепция "эволюционной достаточности" была выдвинута молекулярным биологом Цукеркандлом. Вместе с тем она очень напоминает вывод, сделанный генетиком-популяционистом Ивенсом [1757]. По его мнению, специфические требования окружающей среды могут удовлетворяться за счет различных, не обязательно "оптимальных" комбинаций генов. Идеи Кимуры, развитые им в недавнем обзоре [1941], также довольно близки к этой концепции. Основное различие между взглядами данных исследователей состоит в следующем: по Ивенсу и Цукеркандлу положительный отбор, т. е. отбор слабо благоприятных замен, возможно, имеет несколько большее значение, чем полагает Кимура, рассматривающий его как некое дополнение к случайным процессам. Однако эти авторы согласны в том, что в постоянной среде действует главным образом отбор отрицательного типа, т. е. отбор, стремящийся сохранить определенную функцию путем элиминации вредных мутантов.
Ограниченность современных представлений о естественном отборе и нейтральных заменах при эволюции белков. Как уже отмечалось, большинство исследователей придерживаются мнения, что естественный отбор обусловил замену некоторых аминокислот в белках и существование некоторых систем генетического белкового полиморфизма, выявленных в популяции человека. С другой стороны, часть межвидовой изменчивости и изменчивости внутри популяции человека, вероятно, возникла в результате случайного дрейфа, при этом селективное преимущество или вредность могут быть сравнительно небольшими или даже полностью отсутствовать. Однако имеющиеся в настоящее время данные не позволяют ответить на вопрос о том, какая доля генетической изменчивости обусловлена отбором, а какая - случайными процессами. В этом контексте следует напомнить величину генетического полиморфизма в популяции людей: геном человека, вероятно, содержит около 50000-100000 структурных генов [1943], кодирующих белки. Известно несколько сотен таких генов, причем до 30% из них могут быть полиморфными.
Системы полиморфизма выявлены главным образом при изучении белков крови. Мы уже говорили о том, что полиморфизм белков других, менее удобных для исследования тканей, возможно, намного ниже; однако важно помнить, что полиморфных локусов существует сотни и даже тысячи, а нам из них известна лишь небольшая часть. Кроме того, мы совершенно не знаем физиологической функции многих полиморфных ферментов. Вместе с тем выводы относительно естественного отбора окажутся гораздо правдоподобнее, если они будут основываться на знании физиологической функции изучаемой системы полиморфизма.
Специфическая функция большинства белков решающим образом зависит от немногих аминокислотных позиций. Функциональные ограничения носят столь общий характер, что они вполне совместимы с множеством различных аминокислот; например, трехмерная структура белка может сохраняться при самых разнообразных аминокислотных заменах. При этом в результате генетического дрейфа может происходить сдвиг частот тех или иных оснований, что в свою очередь приводит к возникновению полиморфизма на уровне белков. Системы полиморфизма детерминируют небольшие функциональные различия, не влияющие или лишь незначительно влияющие на приспособленность (разд. 6.2.1.1) их носителей, и вызывают действие естественного отбора. При изменении экологических условий полиморфные системы могут стать источником наследственной изменчивости и обеспечить быструю адаптацию. С другой стороны, тот факт, что для большинства систем полиморфизма селективные влияния пока неизвестны, не означает, что отбор отсутствовал. Просто его трудно обнаружить, особенно среди населения экономически развитых стран, где современная цивилизация значительно изменила условия жизни людей, исключив некоторые потенциально весьма существенные селективные факторы, например инфекционные болезни и недоедание. Для выяснения соответствующих селективных механизмов необходимо сформулировать специальные, обоснованные с функциональной точки зрения гипотезы. Это не означает, что все функциональные различия между полиморфными вариантами когда-то должны были сказаться на приспособленностях. Вместе с тем, не приняв такого предположения, было бы трудно объяснить тот факт, что редкие варианты полиморфных ферментов, как правило, обладают пониженной активностью (разд. 6.1.2). Бесспорно, что для большинства полиморфных систем человека какие-либо селективные влияния в настоящее время неизвестны, однако отсюда нельзя делать вывод об отсутствии отбора; скорее этот факт свидетельствует о нашей неспособности выдвинуть и проверить обоснованные гипотезы относительно селективных механизмов. При изучении полиморфизма человека гипотеза нейтральности, возможно, играет даже отрицательную роль, поскольку она не нацеливает на выявление факторов, осуществляющих естественный отбор.
Молекулярные часы эволюции и мутации. Как уже отмечалось, существование эволюционных часов можно объяснить в том случае, если накопление мутаций зависит от времени и не зависит от вида организма и если замены фиксируются в результате случайных процессов. Из разд. 5.1.3 мы узнали, что частоты возникновения некоторых мутаций человека для мужских половых клеток выше, чем для женских, что частоты возникновения ряда мутаций увеличиваются с возрастом отцов и что многие мутации, вероятно, связаны с репликацией ДНК. Существование сильных различий в продолжительности поколений разных животных делает гипотезу о простой зависимости накопления мутаций от времени весьма маловероятной.
Для получения более реальной величины скорости мутирования было бы желательно составить для разных видов соответствующее "расписание", основанное на числе циклов репликации ДНК в единицу времени, однако мы не располагаем необходимой информацией о кинетике деления половых клеток [1985]. Если бы в замене оснований существовала регулярность, подобная той, что свойственна ходу часов (в чем мы сомневаемся), это определенно свидетельствовало бы против случайной фиксации мутаций: ведь по Кимуре скорость (частота) фиксации зависит только от частоты возникновения мутаций.
Выйти из затруднительного положения можно следующим образом. Можно предположить, что мутации, слабо вредные в больших популяциях, в малых популяциях нейтральны, и поэтому вероятность их фиксации на самом деле выше: виды с большими размерами тела (например, слоны) как правило имеют значительно большее время инерции (и, вероятно, более низкое "число циклов репликации ДНК в единицу времени"), но меньшую популяционную численность, чем виды с небольшими размерами тела (например, мыши). Можно привести и другой аргумент, например, что "частота мутаций в единицу времени", не только зависит от "числа циклов репликации в единицу времени", но что в результате естественного отбора она приблизилась к некоей оптимальной величине. Непонятно только, как это могло произойти, если большинство мутаций так или иначе нейтральны?
Предлагая свою "теорию нейтральности", Кимура выдвинул, несомненно, важное положение, подчеркнув, что эволюция на молекулярном уровне имеет некоторые особенности, не выявляемые при изучении эволюции на уровне фенотипов. Почти нет сомнений, что случайные (или почти случайные) процессы на молекулярном уровне играют гораздо большую роль, чем это думали большинство биологов. Создатели новых теоретических концепций часто склонны переоценивать их объясняющие возможности. Однако, как утверждает Поппер, наука может развиваться только путем смелого выдвижения гипотез и последующей строгой их проверки.
Эволюция путем перетасовки экзонов. Открытие экзон-интронной структуры генов (разд. 2.3 и 4.3) расширило наши представления об эволюции белков: экзоны могут разделяться и выстраиваться в каком-то новом порядке, у одного вида могут экспрессироваться только некоторые экзоны данного гена, а у другого - весь их набор. Как уже отмечалось (разд. 4.2.2.4), такие различия в использовании экзонов одного гена наблюдались даже между разными тканями одних и тех же особей; эти различия в транскрипции генов могут служить одним из механизмов дифференцировки.
Некоторые наборы экзонов используются при сборке разных белков. Так, например, рецептор липопротеинов низкой плотности обнаруживает гомологию с восемью экзонами, кодирующими молекулу предшественника фактора роста эпидермиса. Этот и другие результаты свидетельствуют о том, что функциональные белки представляют собой мозаику из более простых структур, подвергающихся перетасовке [1925а]. Сложную структуру белков можно объяснить комбинированием сравнительно небольшого числа маленьких генов, определяющих структуру экзонов.
Сравнение данных по белкам с данными по хромосомам и сателлитной ДНК. Данные об эволюции белков свидетельствуют, что различия между белками Homo и таких высших приматов, как шимпанзе и горилла, удивительно малы. Можно считать, что эти белки практически одинаковы. Например, видовые различия молекул гемоглобина с функциональной точки зрения менее значительны, чем различия между редкими вариантами, имеющимися в популяциях человека, которые, хотя и могут приводить к легкой гемолитической анемии, вполне совместимы с жизнью. Такую крайне медленную эволюцию можно объяснить, предположив, что функция этих белков осталась в основном неизменной. Если мы обратимся к кариотипам, то обнаружим, что они отличаются небольшим числом хромосомных перестроек, главным образом перицентрических инверсий. Похожие перестройки встречаются, причем не так уж редко, в современной популяции человека и совсем не влияют на фенотип. Ими можно было бы объяснить образование репродуктивных барьеров, бывших когда-то важным условием видообразования; однако они ничего не говорят нам о генетических механизмах, обусловивших формирование специфического фенотипа человека. О функциях добавочных R- и Т-сегментов и о видовых различиях по гетерохроматиновому материалу и сателлитной ДНК известно мало. Установлено, что прицентромерный гетерохроматин проявляет высокую изменчивость в современных популяциях человека. Была выдвинута гипотеза о возможном влиянии этих гетероморфизмов на фенотипы человека, например поведенческие, однако признания среди специалистов она не получила [1914].
Нам остается сделать вывод, что гены, важные для эволюции человека в течение периода, когда происходило преобразование его мозга, совершенно неизвестны. Поскольку большая часть ДНК человека не кодирует белков и либо вообще не нужна, либо участвует в регуляции генной активности (разд. 4.8), можно предположить, что соответствующие изменения локализованы именно в этой, не содержащей структурных генов ДНК [1993]. Такие изменения могли произойти в неэкспрессируемых участках ДНК, относительно которых постулируется, что они имеют регуляторные функции. Возможно, что нуклеотидные последовательности ДНК, несущественные для реализации функций структурных генов, необходимы для развития, и, следовательно, изменения таких последовательностей могли оказать особое влияние на преобразования функции мозга. Однако эта идея весьма спекулятивна и носит слишком общий характер. Чтобы сформулировать более конкретные гипотезы, необходимо больше знать о генетической детерминации эмбрионального развития и о генах, влияющих на межвидовую изменчивость поведенческих признаков (гл. 8). Даже если исключить из рассмотрения все фенотипические эффекты и ограничиться анализом таких известных генетических феноменов, как хромосомные перестройки, добавление или потеря материала хромосом, изменчивость сателлитной ДНК и аминокислотных последовательностей белков, все равно придется констатировать слабое понимание многих аспектов эволюционного процесса. Например, мы не знаем, как происходит фиксация хромосомных перестроек в популяциях. Идентичны ли механизмы их фиксации тем процессам, которые приводят к фиксации аминокислотных замен? Какие элементарные события привели к образованию разных типов сателлитной ДНК? Можно ли говорить о специфическом значении таких событий для видообразования или для изменений в регуляции функций генов?
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'