XVI. Взаимодействие генов
Многие гены действуют совместно (или, иначе говоря, взаимодействуют) при формировании признака. Мы уже познакомились с такими примерами в предыдущих главах.
Чалая масть крупного рогатого скота выглядит по-разному в зависимости от фона: рыжего или черного. Слепоту или глухоту могут вызывать многие аномальные гены (см. главу IX), поскольку способность видеть или слышать определяется взаимодействием многих генов. В настоящей главе мы рассмотрим несколько случаев взаимодействия генов более подробно.
Оперение у попугайчика-неразлучника
Дикий тип попугайчика-неразлучника имеет ярко-зеленое оперение. Мы не знаем, да и не можем знать, взаимодействием скольких генов определяется эта окраска, поскольку ген обнаруживается только в случае его существования по меньшей мере в двух различных аллельных формах. Для некоторых генов, контролирующих цвет оперения, известны различные аллели. Рецессивный ген у определяет желтый цвет перьев, другой рецессивный ген (b) - голубой. Нормальные аллели обоих генов (Y и В) должны быть в числе многих генов, взаимодействие которых вызывает зеленую окраску оперения.
Скрещивание между голубыми и желтыми птицами должно быть записано следующим образом:
Р1 bb YY + ВВ yy (см. первое правило в главе XIV). F1 Вb Yy
Поскольку гены b и y рецессивны, птицы F1 имеют обычное зеленое оперение. Это явление аналогично рождению нормальных детей у глухих родителей, если их глухота обусловлена двумя разными рецессивными генами (см. главу IX).
Скрещивание между птицами F1 можно записать так:
В результате птицы F2 будут следующими:
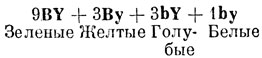
Таким образом, взаимодействие генов, определяющих голубую и желтую окраску у птиц с генотипом bb yy, вызывает белое оперение. Это может показаться удивительным, так как смесь голубого и желтого дает зеленый цвет. Но рассмотрим это с другой точки зрения. Зеленая окраска диких птиц вызывается смешиванием голубого и желтого цветов. Птицы с генотипом bb голубые, так как в их оперении нет желтого цвета. Птицы с генотипом yy желтые, так как в их оперении нет голубого цвета. Птицы с генотипом bb yy не имеют в оперении ни желтого, ни голубого цвета, и поэтому они не окрашены, т. е имеют белое оперение.
В этом примере взаимодействие двух разных генов вызывает появление нового признака, который не может быть обусловлен действием какого-либо одного гена.
Окраска шерсти у мыши
Начнем с повторения того, что нам уже известно об этом признаке (см. главу X) Мышь дикого типа (или агути) имеет черную шерсть с желтыми перетяжками. Это и приводит к тому, что шерсть выглядит пестрой Рецессивный ген а (неагути) определяет отсутствие желтой перетяжки. Поэтому животные с генотипом аа совершенно черные.
Другой несцепленный ген 6 заменяет черную окраску коричневой. Этот ген также рецессивен и коричневый пигмент образуется только у гомозиготных мышей (bb), Окраска шерсти таких мышей зависит от того, являются ли они агути или неагути. Если мышь bb - агути, то каждый ее волосок имеет желтую перетяжку, и шерсть оказывается пестро-коричневой. Эту окраску называют коричной. Если мышь bb - агути, желтые перетяжки отсутствуют и шерсть имеет ровную коричневую окраску. (Таких мышей называют шоколадными.) И в этом случае взаимодействие двух различных рецессивных генов вызывает появление нового фенотипа.
Если обозначить нормальные аллели генов а и b буквами А и В, то можно составить следующую таблицу (табл. 15):

Таблица 15
На основании этих данных легко предсказать результаты скрещивания между черными и коричными мышами:

Многие другие гены влияют на окраску шерсти, и их взаимодействие создает большое разнообразие окрасок, известных любителям. Ген, определяющий альбинизм (с), при этом играет особую роль, так как у мышей, гомозиготных по гену с, шерсть лишена всякой окраски. Такие мыши - всегда альбиносы (сс) независимо от остальных генов окраски шерсти, которые они несут. Поэтому при использовании в скрещиваниях альбиносов неизвестного генотипа можно получить самое неожиданное потомство.
Поскольку ген альбинизма, кроме того, определяет отсутствие окраски радужной оболочки глаз, альбиносы имеют розовые глаза. Альбиносы среди людей имеют белую кожу, белые волосы и розовые глаза независимо от того, какие у них гены окраски кожи, глаз и волос. Предположим, мальчик покупает мышей - самца шоколадной окраски и самку-альбиноса - в зоомагазине. Он скрещивает их и получает помет, состоящий целиком из мышат агути. Если мальчик немного знаком с генетикой, то такой результат скрещивания покажет ему, что самка-альбинос несет нормальные аллели генов а и b. Это скрещивание поэтому может быть записано так:

Какое потомство можно ожидать от скрещивания между мышами поколения FA? На этот вопрос можно ответить, перемножив
С другой стороны, можно воспользоваться тем, что нам уже известно о расщеплении по двум первым парам генов (А-а и В-b). Третья пара (С-с) не сцеплена ни с одной из этих пар генов и поэтому расщепляется независимо от них. В соответствии с первым законом Менделя скрещивание между особями с генотипом Сс дает в потомстве 3/4 окрашенных мышей и 1/4 альбиносов. Таким образом, в каждом из четырех типов мышей, образующихся в результате расщепления по генам А-а и В-b, будет 3/4 окрашенных и 1/4 мышей альбиносов. Это определяет следующее окончательное соотношение особей F2:

Если вы вернетесь к стр. 108, то увидите, что это то же отношение (27:9:9:9:3:3:3:1), которое было получено и при скрещивании особей крупного рогатого скота, различающихся по трем парам несцепленных генов. В данном случае все животные-альбиносы выглядят одинаково, в связи с чем отношение изменяется следующим образом: 27 агути: 9 коричных : 9 черных : 3 шоколадных : 16 альбиносов.
При суммировании всех окрашенных животных отношение их числа к числу альбиносов оказывается равным 48:16 = 3:1, что и следовало ожидать на основании первого закона Менделя Это отношение всегда можно обнаружить в F2 при скрещивании (P1) альбиноса и окрашенной мыши, если только сама окрашенная мышь не несет гена, вызывающего альбинизм. Но какой именно тип (или типы) окраски шерсти будет у потомства, обычно не удается предугадать.
На этих примерах мы видели, как взаимодействие двух генов приводит к появлению нового фенотипа (шоколадных мышей). Кроме того, мы познакомились с новым типом взаимодействия генов на примере гена альбинизма, который своим действием подавляет проявление других генов.
Взаимодействие генов у человека
Рост индивидуума отчасти определяется его генотипом, а отчасти внешней средой, в которой он рос. В главе XVIII мы еще вернемся к вопросу о влиянии внешней среды на размеры тела. В настоящей главе рассмотрим гены, контролирующие только рост человека. Некоторые из этих генов вызывают появление совершенно ненормальных признаков, таких, как карликовость или гигантизм. В определении нормального роста решающую роль, очевидно, играют нормальные аллели этих генов. Но даже совершенно нормальные люди заметно отличаются по росту, что в большей части обусловлено их генотипами. Отдельные гены, вызывающие изменчивость роста в пределах нормы, оказывают небольшое действие: некоторые гены несколько увеличивают рост, другие - его уменьшают, Совместное действие всех этих генов и определяет рост человека в той мере, в какой он зависит от генотипа. Гены, действующие таким образом, называются полимерными генами, а признаки, которые они контролируют, - мерными, или количественными признаками, так как различия между ними носят количественный, а не качественный характер (см. главу IX). Размеры тела и умственные способности человека, молочная продуктивность коров, яйценоскость кур - это примеры количественных признаков, которые контролируются полимерными генами, а также подвергаются влиянию окружающих условий.
Количественные признаки не дают четких менделевских отношений. Несомненно, каждый из многочисленных полимерных генов, контролирующих, например, рост человека, передается по правилам, установленным Менделем, и дал бы менделевские отношения, если бы можно было отделить его действие от действия других генов и окружающих условий.
Но взаимодействие между генами и между генами и внешней средой определяет непрерывный ряд индивидуумов от самого высокого роста до самого низкого, в котором нельзя различить никаких четких классов.
Мы сможем представить, каким образом наследуются количественные признаки, если рассмотрим вымышленный пример, который чрезвычайно упрощает реальное положение.
Забудем на время о влиянии внешней среды и допустим, что существует всего три пары генов, контролирующих рост человека, и что все они не сцеплены. Обозначим эти гены буквами А и а, В и b, С и с. Индивидуум с генотипом аа bb cc имеют самый низкий рост - 150 см. Допустим также, что каждый доминантный ген добавляет к росту 5 см, так что, например, человек с генотипом аа Вb Сс имеет рост 160 см, а человек с генотипом АА bb СС - 170 см. Люди с генотипом АА ВВ СС самые высокие: их рост равен 180 см.
Теперь представим, что племя людей низкого роста (аа bb сс) порабощается ордой воинов из племени, состоящего только из высоких людей (АА ВВ СС). Победители убивают мужчин и женятся на женщинах порабощенного племени. Как же распределятся их дети и внуки по росту?
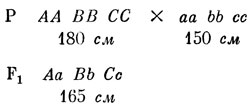
Таким образом, все дети будут иметь рост, промежуточный между ростом родителей. Что можно сказать о внуках? Поскольку все дети имеют одинаковый генотип, браки между ними можно записать так:
F2 можно охарактеризовать, составив решетку из 64 квадратов (8 × 8). Но мы снова воспользуемся более быстрым способом перемножения. В данном случае индивидуум, гомозиготный по доминантному гену, будет выше гетерозиготного, и расщепление приведет к соотношению фенотипов не 3:1, наблюдающемуся при полном доминировании, а к отношению 1:2:1, имеющему место в случае неполного доминирования (см. главу V). Результат расщепления, например, по А и а должен быть записан так: 1АА + 2Аа + 1aa. Таким же образом можно записать результаты расщепления и по двум остальным парам генов. Комбинация результатов всех трех расщеплений в соответствии со вторым законом Менделя может быть представлена в виде
Не рассматривая конкретно все получающиеся генотипы, можно рассчитать, сколько генотипов будут иметь 6, 5, 4, 3, 2, 1 или 0 доминантных генов, так как именно это и позволит нам установить распределение индивидуумов по росту. В табл. 16 показан расчет для 6, 5, 4 и 3 доминантных генов.
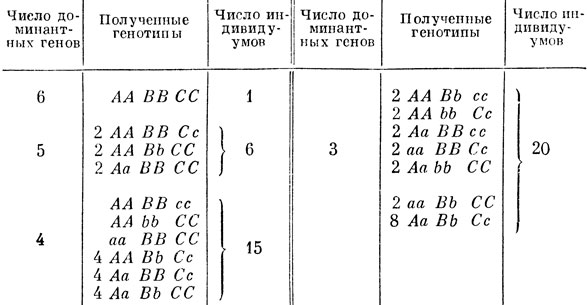
Таблица 16
Индивидуумы с 2, 1 и 0 доминантных генов имеют соответственно 4, 5 и 6 рецессивных, так как у каждого организма должно быть всего 6 генов. Число таких индивидуумов получить очень легко, если в соответствующей части приведенной таблицы заменить каждую заглавную букву маленькой и каждую маленькую - заглавной (например, 2АА ВВСс во второй строчке заменить 2 аа bb сС). Это показывает, что особей с 4, 5 и 6 рецессивными генами столько же, сколько и с соответствующим числом доминантных генов: 15, 6 и 1.
Теперь нам известно все, что нужно для установления распределения людей по росту в F2 (табл. 17).

Таблица 17
На рис. 37 эти результаты представлены графически. Колонка, следующая за каждой из фигур, показывает, какую часть F2 составляют индивидуумы данного роста. Вы видите, что люди среднего роста самые многочисленные, а очень низкие и очень высокие встречаются редко.
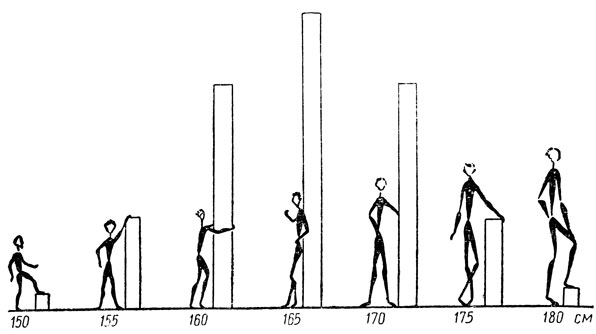
Рис. 37. Относительная частота появления индивидуумов разного роста при условии, что все различия в росте обусловлены тремя парами генов, подчиняющихся закону свободных сочетаний
Как я уже говорила, рассмотренный пример дает чрезвычайно упрощенную картину наследования размера тела. На самом деле этот признак определяется не тремя, а значительно большим числом пар генов. Участие отдельных генов в определении роста очень невелико и не обязательно одинаково. Некоторые гены могут иметь некоторую степень доминирования. Если бы мы рассмотрели все пары генов, определяющие рост, то получили бы значительно большее число групп и уменьшили бы тем самым различия между отдельными классами роста. Если бы, кроме того, мы учли, что индивидуумы даже с одинаковыми генотипами могут различаться по росту в зависимости от питания или других факторов внешней среды, то классы вообще бы не удалось выделить, и график выглядел бы так, как показано на рис. 38.
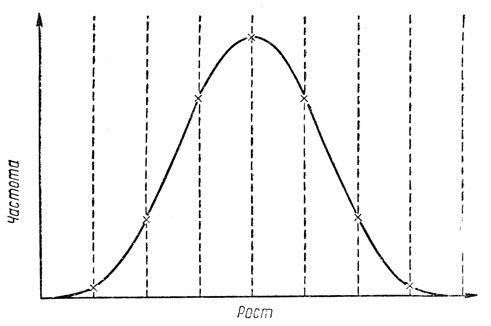
Рис. 38. Распределение частоты появления индивидуумов разного роста при учете действия всех генов, контролирующих рост, и влияния внешней среды
Этот вид распределения характерен для количественных признаков и, в частности, для умственных способностей (за исключением случаев слабоумия, которое, как и карликовость, появляется в результате резких нарушений в развитии, нередко вызываемых аномальными генами) Среди нормальных людей чаще всего встречаются индивидуумы со средними умственными способностями, в то время как очень глупые или, наоборот, одаренные встречаются редко. Между средними и крайними выражениями этого признака мы находим все промежуточные; умственные способности, приближающиеся к средним, встречаются чаще, чем приближающиеся к крайним.
Выводы
1. Формирование данного признака осуществляется взаимодействием многих генов. В настоящей главе мы рассмотрели примеры, касающиеся следующих типов взаимодействия генов:
а) появление нового фенотипа в результате взаимодействия нескольких пар генов; например, среди попугайчиков-неразлучников гомозиготы по голубой и желтой окраске имеют белое оперение; среди мышей гомозиготы по окраске неагути и по коричневой окраске имеют сплошную коричневую (шоколадную) шерсть;
б) подавление действия одного гена действием других генов. Например, в организмах, гомозиготных по гену альбинизма, подавляется действие всех генов, определяющих цвет волос, кожи и глаз.
2. Взаимодействие генов может изменять или скрывать ожидаемые менделевские отношения. По признаку, определяемому многими полимерными генами, не происходит расщепления на четко различимые классы фенотипов. Распределение индивидуумов в потомстве характеризуется постепенными переходами от одного фенотипа к другому; при этом большинство индивидуумов имеют средние показатели, в то время как особи с крайними показателями встречаются редко.
Задание
1. Кролик альбинос (♂) скрещен с самкой неагути (черная). В помете было пять крольчат агути. Какие генотипы имели родители, крольчата? Какие фенотипы вы ожидаете встретить в F2?
2. У душистого горошка имеются две разновидности с белыми цветками, У каждой разновидности белая окраска обусловлена рецессивным геном. Если скрестить растения этих разновидностей, то все потомство F1 оказывается с окрашенными цветками, но в F2 снова появляются растения с белыми цветками. Как вы это объясните? Какая часть растений F2 имеет белые цветки?
3. Проведено скрещивание двух сортов кукурузы У растений одного сорта средняя длина початка составляла 6-7 см, а у растений другого сорта - 16-17 см. Растения F1 имели промежуточную длину початков. В F2 было получено следующее распределение растений:

Изобразите графически распределение растений F2, как это сделано на рис. 38.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'