III.2.2. Нарушение нормальной конъюгации гомологичных хромосом и негомологичное спаривание - одна из причин равновероятного образования гамет разного генотипа
Основой для изучения влияния разных событий в мейозе (конъюгации, кроссинговера и расхождения хромосом) являются эксперименты, в которых обнаруживаются систематические нарушения расщеплений, сопровождающиеся увеличением стерильности, появлением анеуплоидов и т. п.
До недавнего времени считалось, что расхождение хромосом определяется кроссинговером, обусловленным конъюгацией гомологов. Для выяснения связи между процессами конъюгации, кроссинговера и расхождения хромосом Чадовым и сотр. были проведены исследования, показавшие, что частичный асинапсис гомологов приводит к спариванию и нерасхождению негомологичных хромосом, что свидетельствует о независимости расхождения хромосом от кроссинговера. Эти заключения сделаны на основе гибридологического анализа без цитологических наблюдений (Чадов и др., 1981, 1985, 1986).
В опытах использовали самок дрозофилы, имевших нормальную метацентрическую хромосому 2 и два акроцентрика, каждый из которых содержал одно из структурно-нормальных плеч второй аутосомы - левое - F(2L) или правое - F(2R), центромеру 2 и участок прицентромерного гетерохроматина противоположного плеча (линия МАК-2) (рис. III. 1). Поскольку спариваться могут только две хромосомы, наличие трех центромер приводит к асинапсу прицентромерной области правого или левого акроцентрика. Было показано, что асинапсис прицентромерного района вызывает нерасхождение метацентрика с одним из акроцентриков (или двух акроцентриков) и образование анеуплоидных гамет. Для оценки частоты возникновения анеуплоидных гамет у самок МАК-2, маркированных рецессивной мутацией а* по левому акроцентрику (генотип 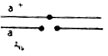 ), их скрещивали с самцами четырех генотипов: 1 - с нормальной второй хромосомой, маркированной мутацией а в левом и с - в правом плече (генотип
), их скрещивали с самцами четырех генотипов: 1 - с нормальной второй хромосомой, маркированной мутацией а в левом и с - в правом плече (генотип 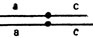 ); 2-c моноцентрической хромосомой, у которой левое плечо второй хромосомы представлено дважды (так называемая компаунд-хромосома - C(2L)**. Она маркирована рецессивной мутацией d и двумя правыми плечами второй хромосомы - F(2R). Генотип
); 2-c моноцентрической хромосомой, у которой левое плечо второй хромосомы представлено дважды (так называемая компаунд-хромосома - C(2L)**. Она маркирована рецессивной мутацией d и двумя правыми плечами второй хромосомы - F(2R). Генотип 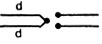 - C(2L), d; F(2R); 3 - с компаунд-хромосомой по правому плечу, маркированному мутацией с, и двумя левыми плечами с геном а - генотип
- C(2L), d; F(2R); 3 - с компаунд-хромосомой по правому плечу, маркированному мутацией с, и двумя левыми плечами с геном а - генотип 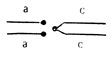 -F(2L), а\ C(2R), с\ 4 - с двумя компаунд-хромосомами - левой с геном d и правой с геном b - генотип
-F(2L), а\ C(2R), с\ 4 - с двумя компаунд-хромосомами - левой с геном d и правой с геном b - генотип 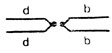 -C(2L), d; C(2R), b. В каждом скрещивании анализировали расщепление по соответствующему маркеру: в 1-ом - по а, во 2-м - по d, в 3-м - по с и в 4-ом - по b.
-C(2L), d; C(2R), b. В каждом скрещивании анализировали расщепление по соответствующему маркеру: в 1-ом - по а, во 2-м - по d, в 3-м - по с и в 4-ом - по b.
* (Обозначения генов-маркеров - а, b, с и d - условные.)
** (Большой практикум по генетике животных и растений. М., 1977. С. 111-118)
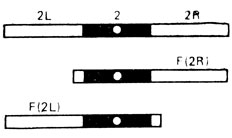
Рис. III. 1. Структура линии МАК-2 у дрозофилы. Черным цветом обозначен прицентромерный гетерохроматин. Левое плечо F (2L) терминировано теломерным участком 2L, а правое - F (2R) - теломерным участком 2R. В гетерозиготе - 2/F (2L); F (2R) - центромера с окружающим ее гетерохроматином представлена трижды
Очевидно, что и у самок МАК-2 и у самцов второй, третьей и четвертой линий вследствие особенностей их генетической структуры могут образовываться гаметы, анеуплоидные по второй хромосоме, большинство из которых оказываются зиготическими леталями. Лишь часть из них жизнеспособны, их можно выявить по маркерам - b, с, d и по частоте их выщепления определить частоту нерасхождения хромосом у самок МАК-2 (табл. III. 1).
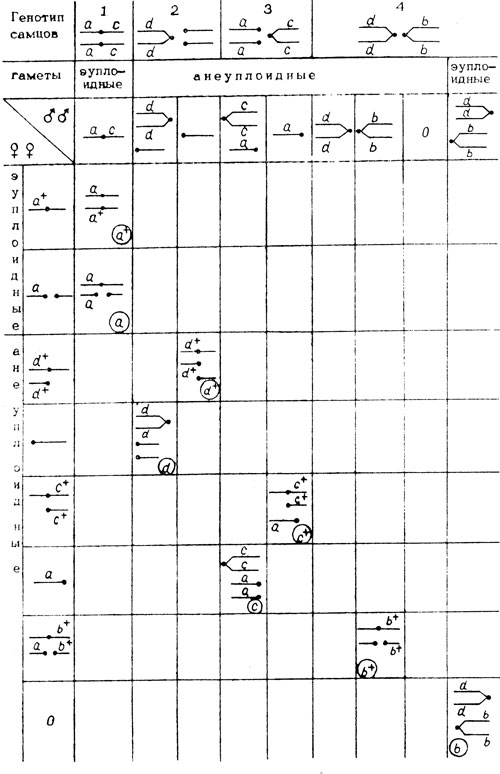
Таблица III. 1. Теоретически ожидаемые классы, возникающие с участием эу- и анеуплоидных гамет, в потомстве самок МАК-2 при скрещивании их с самцами четырех разных генотипов
Частоту оценивали по отношению числа эу- и анеуплоидных гамет разного типа к общему числу яиц, отложенных самкой в данном скрещивании. Оказалось, что общая частота жизнеспособных гамет у самок МАК-2 составляла примерно 90%, из них - 66,1% эуплоидных и 24,2%-анеуплоидных. Анеуплоидных потомков практически не было - они погибали, не достигая стадии имаго; часть яйцеклеток отмирала из-за дистрофических явлений (табл. III. 2).

Таблица III.2 Соотношение фенотипов по маркерам а, b, с и d, генотипы гамет самок МАК-2 и их частоты в скрещиваниях с самцами 1-, 2-, 3- и 4-ой линий
Для объяснения полученных данных были рассмотрены 3 варианта возможных расхождений хромосом 2 у самок МАК-2 при разных типах их ориентации в мейозе и определен теоретически ожидаемый процент эу- и анеуплоидных гамет (рис. III. 2).
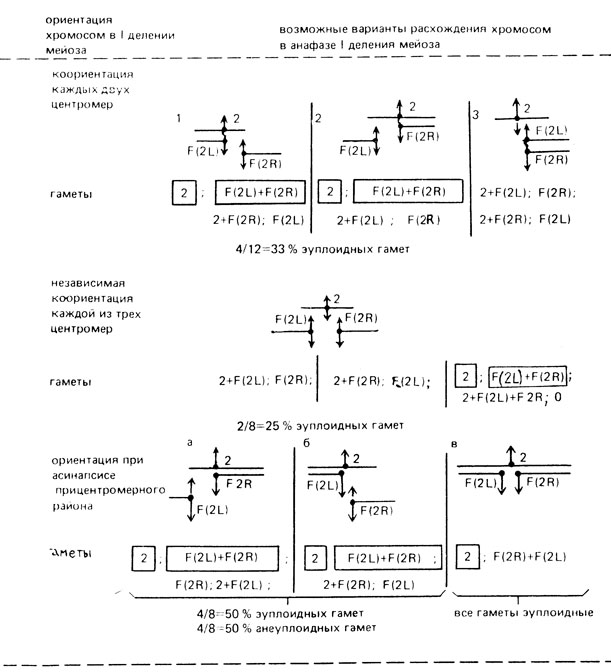
Рис. III.2. Ожидаемые типы гамет и их соотношение при разных вариантах ориентации хромосом 2 у самок МАК-2. В рамках - эуплоидные гаметы. Стрелки показывают направление к полюсам
При рассмотрении этих вариантов становится очевидным, что частота эуплоидных гамет, полученная в опыте (66,2%), не может быть объяснена коориентацией двух центромер или независимой коориентацией каждой из трех центромер, так как в этих случаях ожидается возникновение 33% или 25% эуплоидных гамет соответственно.
Следовательно, в основе возникновения эуплоидных гамет у самок МАК-2 лежит асинапсис прицентромерного района, приводящий к образованию либо равных количеств эу- и анеуплоидных гамет, либо только эуплоидных гамет (рис. III. 2, а, б, в). Исходя из этого, можно определить частоту эуплоидных гамет у самок МАК-2 по доле анеуплоидных гамет. Если бы мейоз у них проходил только по типу а и б (рис. III.2), то доля эуплоидных гамет была бы такая же, как и анеуплоидных, т. е. 24,2%. В опыте она -66,2%, тогда 42% (66,2-24,2) эуплоидных гамет возникает при расхождении по варианту в (Чадов, 1986а).
Таким образом, асинапсис прицентромерного района ведет к нерасхождению хромосом и отбору на уровне гамет: анеуплоидные гаметы лишь частично принимают участие в оплодотворении. Было установлено также, что у самок МАК-2 асинапсис прицентромерного района второй хромосомы часто приводит к конъюгации любого из акроцентриков - F(2L) или F(2R) - с негомологичными хромосомами, в частности с Х-хромосомой, причем негомологичное спаривание является основной причиной нерасхождения Х-хромосом и нарушения расщепления по генам, локализованным в Х-хромосоме. Это было показано при скрещивании самок МАК-2, у которых Х-хромосома содержала инверсию и была маркирована рецессивной аллелью гена yellow -у- (желтое тело) и доминантной аллелью гена Bar -В- (полосковидные глаза). Самцы имели серое тело и круглые глаза (дикий тип). В норме в F1 такого скрещивания ожидается появление самок с серой окраской тела и глазами, более узкими, чем у нормальных мух, и самцов - желтых с узкими глазами. Появление исключительных самок - с желтым телом и узкими глазами с генотипом 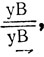 и исключительных самцов - серых с нормальными глазами с генотипом
и исключительных самцов - серых с нормальными глазами с генотипом 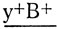 свидетельствовало о нерасхождении Х-хромосом. Частота нерасхождения в этих опытах составляла 15% (Чадов и др., 1986а).
свидетельствовало о нерасхождении Х-хромосом. Частота нерасхождения в этих опытах составляла 15% (Чадов и др., 1986а).
Таким образом, изучение сегрегации хромосом у структурно измененных самок МАК-2 послужило основой для выяснения одного из возможных механизмов нерасхождения хромосом, приводящего к нарушению нормального менделевского наследования.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'