III.2.8. Зависимость расщепления от выживаемости зигот разного генотипа
Неодинаковая жизнеспособность зигот разного генотипа может быть связана с доминантными или рецессивными летальными мутациями, возникающими в гаметах родительских особей. Они могут реализовываться на разных стадиях эмбриогенеза или в постэмбриональном периоде. Естественно, что гибель части зигот влияет на характер расщепления.
Влияние на расщепление доминантных генов с рецессивным летальным действием
Доминантные гены с рецессивным летальным действием обладают плейотропным эффектом; с одной стороны, они определяют доминантный характер проявления какого-либо признака в гете-розиготе, с другой - вызывают гибель особей в гомозиготе, т. е. проявляют свое летальное действие как рецессив. Такие гены известны у многих животных - желтая окраска мышей, корот-коногость у кур, линейная чешуйчатость у карпов, платиновая окраска у лис, окраска ширази у овец и многие другие. Они вызывают отклонение от расщепления 3:1, превращая его в расщепление 2:1. Анализ расщеплений с участием доминантных генов с рецессивным летальным действием осложняется тем, что на сравнительно небольших выборках расщепление 3:1 и 2:1 не всегда возможно различить, а статистический метод проверки по критерию χ2 не позволяет делать выбора из гипотез. Например, если при скрещивании желтых мышей между собой получится расщепление 65 желтых: 32 черных, по методу %2 не отвергаются две гипотезы: 1 - расщепление в опыте соответствует расщеплению 3:1, χ2 = 3,29, p>0,05; 2 - расщепление в опыте соответствует расщеплению 2:1, χ2 = 0,17, p>0,05.
Только на больших выборках расщепление 2:1 отличимо от расщепления 3:1. При суммировании данных по наследованию желтой окраски у мышей, полученных разными исследователями, расщепление при скрещивании желтых мышей между собой было 2386 желтых: 1235 черных - ∑ = 3621, χ2 = 0,96 при Н0 - 2:1 (при Н0 - 3:1 в этом случае χ2 = 160!).
Очевидно, что разные способы генетической проверки, например постановка анализирующего и возвратного скрещиваний, могут способствовать правильному решению вопроса о наследовании. В рассмотренном выше примере это скрещивания: желтые из F1× черные; желтые из F1× желтые Р. В первом случае должно быть расщепление 1:1, во втором - снова 2:1.
Сложнее выяснить, что отклонение есть и оно обусловлено разной жизнеспособностью зигот в том случае, когда гибель части зигот осуществляется на постэмбриональной стадии. В связи с этим необходимо вести контроль плодовитости и отхода в потомстве, если есть предположение о наличии летальности. Основанием для предположения о летальности является невозможность получения единообразного потомства при разведении в себе особей определенного фенотипа. Примеры анализа даны в задачах № III.2 и III.3.
В потомстве от скрещивания серебристо-соболиных норок между собой в F1 всегда наблюдается расщепление: появляются серебристо-соболиные и коричневые особи.
В одном опыте в нескольких пометах от скрещивания серебристо-соболиных особей было получено 93 серебристо-соболиных и 43 коричневых щенка при средней величине помета 3,66 щенка. От скрещивания серебристо-соболиных с коричневыми норками получили 39 коричневых и 41 серебристо-соболиных. В скрещиваниях коричневых норок между собой получали только коричневых I потомков. Величина помета в двух последних скрещиваниях составляла 4,9-5,2 щенка.
Объяснить расщепление, определить генотипы серебристо-соболиных и коричневых норок.
Поскольку серебристо-соболиные норки при скрещивании между собой и с коричневыми всегда дают расщепление, они очевидно гетерозиготны. Для определения числа генов рассмотрим расщепления в опыте:
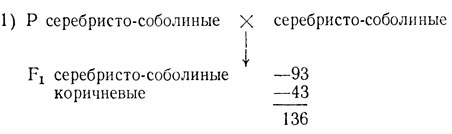
H0 - различия по одному гену, расщепление 3:1, χ2 = 3,2, p>0,05. Отклонение случайно, гипотеза не отвергается.
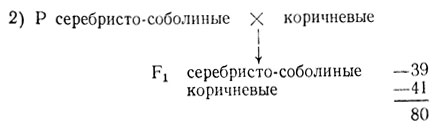
Соотношение хорошо соответствует расщеплению по одному гену 1:1, χ2 = 0,05, p>0,80. Отклонение случайно, гипотеза не отвергается.
Сопоставление данных о плодовитости норок в разных скрещиваниях говорит о частичной гибели зигот при скрещивании серебристо-соболиных норок. Можно предполагать гибель доминантных гомозигот. Тогда генотип серебристо-соболиных норок - Аа, коричневых - аа и расщепление в скрещивании не 3:1, а 2:1 (χ2 = 0,17, p>0,05). Для окончательного утверждения этой гипотезы необходимо поставить дополнительное скрещивание между серебристо-соболиными особями для увеличения выборки и статистической проверки гипотезы 2:1. В некоторых случаях приходится проводить более сложный анализ.
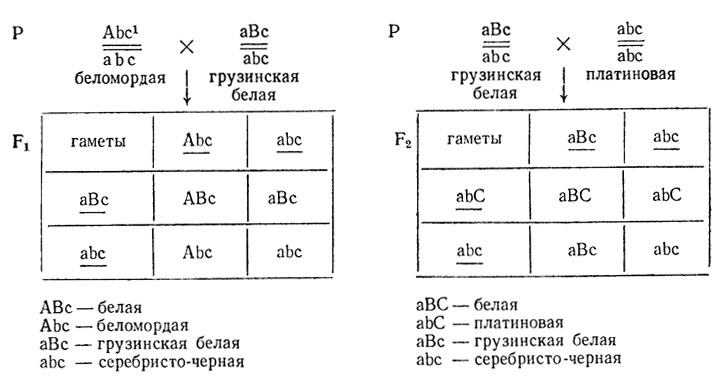
В скрещиваниях платиновых, беломордых и грузинских белых лис с серебристо-черными было установлено, что платиновая, беломордая и грузинская белая окраски, вызывающие общее ослабление пигментации и появление различной пегости, не сцеплены с полом. При скрещивании каждого из этих мутантов с серебристо-черными получали расщепление 2:1, причем серебристо-черная окраска была рецессивной. Следовательно, каждая из них контролируется одним доминантным геном с рецессивным летальным действием. О гибели части мутантов свидетельствуют данные о величине помета: серебристо-черные имеют 4,5 щенка на помет, беломордые 3,5, платиновые и грузинские белые - меньше 3,5.
Для того чтобы установить, затрагивают ли эти мутации один ген или разные, были поставлены скрещивания, результаты которых приведены ниже. Тест на аллелизм в данном случае неприменим, так как мутации доминантные:

Объяснить расщепления, определить генотип всех форм.
1. Расщепления в скрещиваниях 1 и 2 соответствуют соотношению 1:1:1:1 (χ2 = 5,83, p>0,10 для скрещивания 1, χ2 = 0,55, p>0,90 для скрещивания 2). Они могут быть следствием нескольких причин.
1. Поскольку в скрещиваниях 1 и 2 в расщеплении 4 класса и соотношение 1:1:1:1, можно предположить, что исследуемые формы различаются по двум независимо наследуемым доминантным генам с рецессивным летальным действием, взаимодействующим по типу комплементарности. При этом один из генов представлен двумя разными доминантными аллелями.

2. Расщепление 1:1:1:1 может быть следствием тесного сцепления этих двух генов при отсутствии кроссинговера между ними:
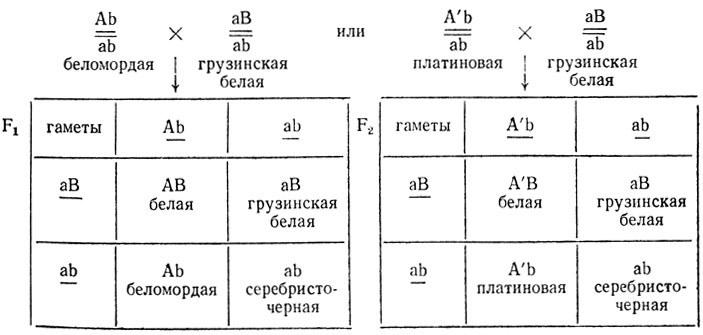
(В решетках Пеннета даны фенотипические радикалы - гены, проявляющие свое действие в фенотипе.)
3. Различия в окраске обусловлены тремя независимо наследуемыми доминантными генами с рецессивным летальным действием. В первом (а) и втором (б) скрещиваниях расщепление происходит по двум разным генам.
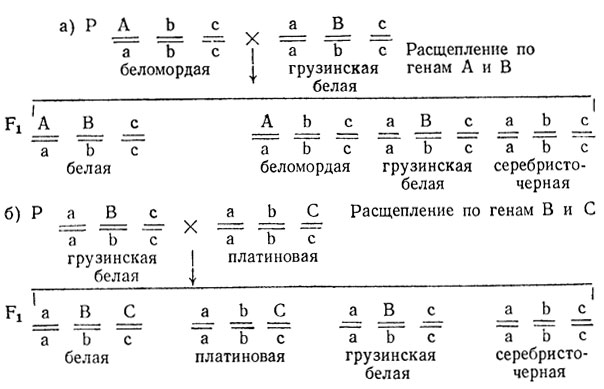
4. Расщепление 1:1:1:1 может быть следствием тесного сцепления трех взаимодействующих генов при отсутствии кроссинговера.
* (Порядок расположения генов может быть иным, он дан произвольно.)
5. Расщепление в соотношении 1:1:1:1 может быть результатом различий по одному гену, представленному серией из четырех аллелей, три из которых - доминантные с рецессивным летальным действием, а четвертый - рецессивный:

Для того, чтобы сделать выбор между этими гипотезами, было поставлено скрещивание - белых с серебристо-черными.
Если бы признак контролировался двумя или тремя генами, то при их независимом наследовании можно ожидать появления четырех фенотипических классов:

Аналогичный результат должен быть в скрещиваниях белых, полученных от платиновой или грузинской белой лис, с серебристо-черными, однако вместо беломордой при этом должны были появиться платиновые, в чем легко можно убедиться, написав соответствующие скрещивания.
Полученное в проверочном скрещивании расщепление - появление беломордых и грузинских белых лис - можно объяснить либо взаимодействием двух (или трех) тесно сцепленных генов, либо взаимодействием трех аллелей при моногенном различии между исходными формами.

Резкое снижение жизнеспособности белых лис, отмеченное в опыте, говорит в пользу действия аллельных мутаций, так как в этом случае белые лисы по генотипу являются компаундами по двум доминантным мутациям одного гена, причем обе - с рецессивным летальным действием. Трудно ожидать снижения жизнеспособности при взаимодействии двух разных генов (комплементарность). Поэтому сделали вывод, что белая, беломордая, грузинская белая, платиновая и серебристо-черная окраски у лис контролируются серией аллелей одного гена, три из которых - доминантные с рецессивным летальным действием. Генотипы лис: белая A1/A2, A1/A3, A2/A3; беломордая A1/а; платиновая A3/а; грузинская белая A2/а; серебристо-черная а/а (по Беляеву и др., 1973).
Следует подчеркнуть, что при множественном аллелизме в случае гетерозиготности исходных форм максимальное число фенотипических классов в расщеплении может быть не 3, а 4, как в случае, описанном выше. В популяции же число возможных генотипов при множественном аллелизме многократно возрастает; оно может быть определено по формуле: 1/2n (n+1), где n - число аллелей. Например, если имеется 7 аллелей какого-либо локуса, число возможных генотипов в популяции будет равно 28: 1/2×7×8 = 28.
Влияние рецессивных летальных мутаций на расщепление
Рецессивные аутосомные и сцепленные с полом летали, вызывая гибель гомозигот в скрещиваниях гетерозигот по летали, могут влиять на расщепление по генам, сцепленным с леталькк При этом соотношение потомков в расщеплении зависит от расстояния между изучаемым геном и леталью, а также от типа гетерозиготы - в цис- или транс-положении гены вводятся в гетерозиготу. Для выявления летали обычно ставятся разные проверочные скрещивания. Примером анализа летали служит задача № III.4.
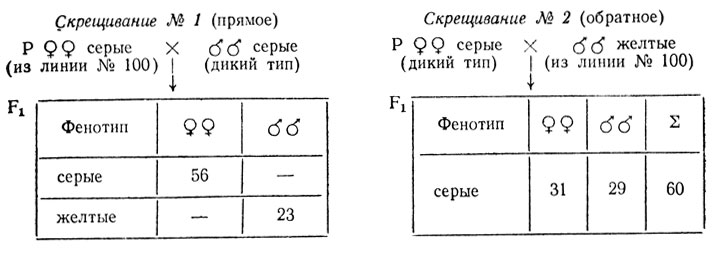
В линии дрозофилы из линии № 100, содержащей инверсии, половина самок имели серую, половина - желтую окраску тела, причем желтые самки оказались стерильными. Все самцы этой линии имели желтую окраску. Соотношение самок и самцов отличалось от нормального, оно больше похоже на расщепление 2♀♀:1♂♂. Было высказано предположение, что недостаток самцов в линии № 100, вероятно, связан с наличием летали в Х-хромосоме серых самок, которые, очевидно, гетерозиготны - в линии происходит расщепление на серых и желтых самок. Для установления генетической структуры этой линии и проверки предположения о наличии летали в Х-хромосоме самок были поставлены скрещивания, результаты которых представлены ниже.
Серые самки из F1 были индивидуально скрещены с желтыми самцами из линии № 100.
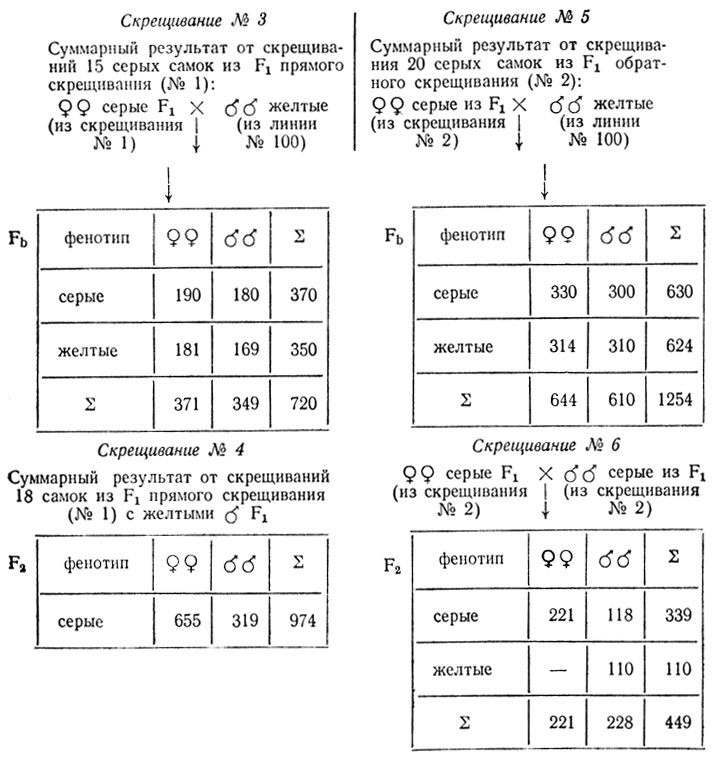
- Серые самки дикого типа (скрещивание № 2) в F1не дают расщепления, что свидетельствует об их гомозиготности, а появление в F2 (скрещивание № 6) серых самок, серых и желтых самцов в соотношении 3/4 серых (♀ и ♂) и 1/4 желтых (♂)свидетельствует о моногенном расщеплении и о доминантности серой окраски (А) над желтой (а), а также о сцеплении признака с полом. Генотипы: серых самок дикого типа
 желтых самцов из линии № 100 -
желтых самцов из линии № 100 - 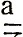 (χ2 = 0,05, p>0,80).
(χ2 = 0,05, p>0,80).
- На основании расщеплений в скрещиваниях № 3 и № 5 (примерно 1/2 серых: 1/2 желтых) можно сделать вывод о моногенном характере различий между исходными мухами и о гетерозиготности самок из F1, использованных в этих скрещиваниях (χ2 = 0,67и 0,92).
- Результаты скрещиваний № 1 и № 4 говорят о гетерозиготности самок из линии № 100, а соотношение полов в потомстве от этих скрещиваний (примерно 2♀:1♀) подтверждают предположение о наличии рецессивной летальной мутации в одной из Х-хромосом серых самок из линии № 100. Очевидно, что эта мутация сцеплена с геном, контролирующим серую окраску (А) тела: в расщеплении уменьшена доля серых самцов. Наличие инверсии в генотипе мух из линии № 100 препятствует прохождению кроссинговера между этими генами.
На основании проведенного анализа запишем схемы всех скрещиваний.
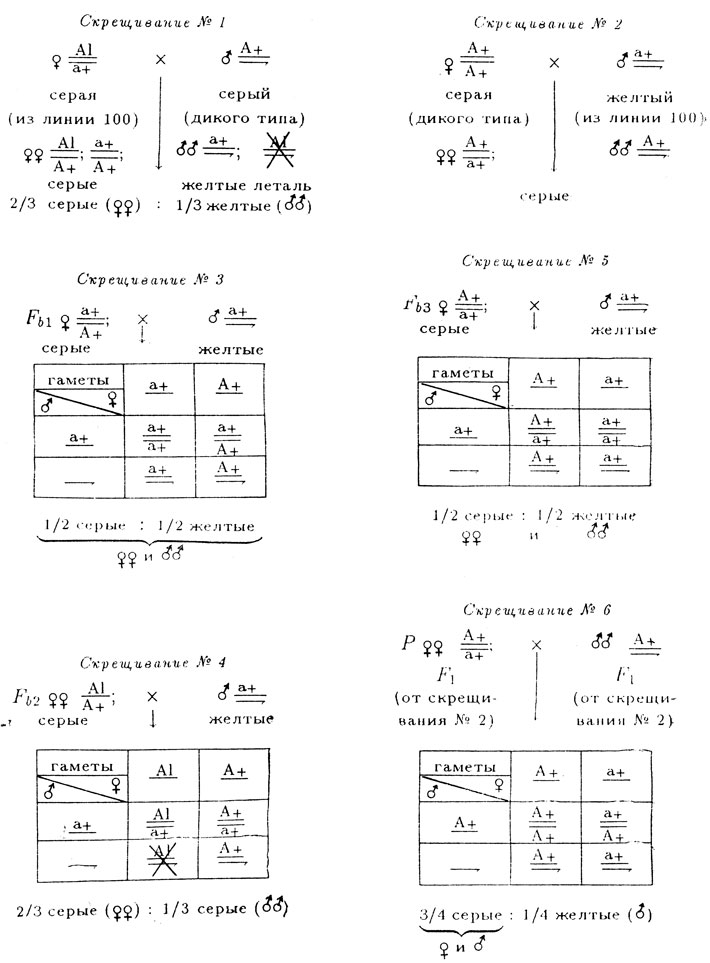
Предложенные гипотезы достаточно хорошо объясняют все результаты. Однако остается невыясненным вопрос о причине стерильности желтых самок в линии № 100. Предлагаем читателю обдумать этот вопрос и предложить какую-либо гипотезу для его объяснения.
У растений многие рецессивные мутации связаны с недостатком или отсутствием хлорофилла, что приводит либо к снижению жизнеспособности растений, либо к их гибели на разных этапах развития. Это служит причиной отклонений в расщеплении, а также вызывает необходимость учета расщеплений не только на проростках, но и на более поздних стадиях развития для определения доли гибели растений и характера наследования признака. Так, у кукурузы гомозиготы по гену wd (white deficiency) имеют белые проростки (расщепление на проростках 3/4 зеленых : 1/4 белых). Однако спустя 1-3 недели происходит отмирание всех белых растений после использования запасов питательных веществ семени и в наиболее поздних стадиях развития растений расщепление исчезает - 3 зеленых : 0 белых. Аналогичные мутации известны у гороха, ячменя, ржи, пшеницы и др.
Другие мутации вызывают гибель лишь части особей на определенном этапе развития, что приводит к уменьшению доли рецессивов в расщеплении и изменению соотношения фенотипов - 4:1, 5:1 и т. п. Эти соотношения варьируют, поскольку, как правило, жизнеспособность таких мутантов в значительной степени зависит от условий.
У человека пониженная жизнеспособность и летальность вследствие действия рецессивных мутаций проявляется в разные периоды эмбриогенеза и на разных этапах развития. Причины снижения жизнеспособности и летального эффекта могут быть связаны как с генными мутациями, так и с хромосомными аномалиями. Цитогенетический анализ абортивных зародышей позволяет установить причину гибели многих из них. В среднем из-за аберраций хромосом на всех сроках беременности происходит более 42% спонтанных абортов: значительная часть новорожденных с хромосомными аберрациями погибает в течение первого и последующих лет жизни.
Среди генных летальных мутаций, приводящих к внутриутробной гибели или смерти в младенческом возрасте, можно назвать рецессивные мутации, вызывающие талассемию, серповидноклеточную анемию, муковисцедоз, врожденный ихтиоз, анэнцефалию (отсутствие мозга), фенилкетонурию и др.
Для изучения летальных или понижающих жизнеспособность мутаций у человека широко используются цитогенетический и биохимический методы анализа, изучение структуры и активности ферментов в норме и при патологии, а также у гетерозиготных носителей; хроматография, разные типы электрофореза.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'