Vl.1.1. Независимое наследование признаков при локализации генов в аутосомах
По результатам расщеплений сначала выясняют наследование по каждому признаку отдельно, затем определяют их совместное наследование.
Если расщепление по каждому признаку происходит согласно менделевским закономерностям, то при независимом наследовании соотношение фенотипов по всем изучаемым признакам определяется путем перемножения формул расщепления, по каждому из них: (3:1)×(3:1); (3:1)×(1:2:1); (3:1)×(9:7); (9:3:4)×(13:3); (3:1)×(12:3:1)×(3:1) и т. д. Поясним это на примерах.
Гибриды F1, полученные от скрещивания растений пшеницы с плотными остистыми колосьями и растений с рыхлыми безостыми колосьями, имели безостые колосья средней плотности, а в F2 произошло расщепление: 58 - безостых с плотным колосом, 62 - безостых с рыхлым колосом, 129 безостых с колосьями средней плотности, 19 остистых с плотным колосом, 43 остистых с колосьями средней плотности, 21 остистых с рыхлым колосом (∑ = 332). Объяснить расщепление, определить, как наследуются признаки, каковы генотипы исходных форм и гибридов F1.
- Анализ по признакам
- Плотность колоса: в F1 единообразие, колосья средней плотности; Р, вероятно, гомозиготны. В F2 расщепление: 77 с плотным, 83 с рыхлым и 171 с колосьями средней плотности. Н0 - расщепление в F2 по одному гену, неполное доминирование, так как в F1 промежуточный тип колоса, а в F2 расщепление примерно 1:2:1; χ2 = 0,66, p>0,70. Отклонение случайно, гипотеза не отвергается. Генотипы: Р - плотный колос - АА, рыхлый колос - аа, F1 - Аа. При неполном доминировании нельзя установить, какой аллель определяет рецессивный признак. Поэтому наши обозначения условны. В данном случае можно было считать рецессивным плотный колос (аа), а рыхлый - гомозиготным по доминантному аллелю (АА).
- Наличие - отсутствие остей. В F1 единообразие, исходные формы, вероятно, гомозиготны. В F2 расщепление: безостые - 248, остистые - 83. Н0 - расщепление по одному гену, 3:1. χ2 = 0,002, p>0,90. Отклонение случайно, гипотеза не отвергается. Генотипы: Р - ВВ - безостый, bb - остистый колос, F1 - Bb - колос средней плотности.
- Анализ совместного наследования признаков В F2 расщепление на 6 фенотипов классов без преобладания родительских комбинаций, Н0 - признаки наследуются независимо. По теории вероятностей расщепление по фенотипу должно происходить по формуле - (1АА:2Аа:1аа)×(3В:1b) = 3ААВ:6АаВ:3ааВ:1AAbb:2Aabb:1aabb, χ2 = 0,65, p>0,98. Отклонение случайно, гипотеза не отвергается.
- Различие исходных растений по плотности колоса обусловлено одним геном. Доминирование неполное - гетерозиготы имеют колос средней плотности.
- Различие исходных растений по наличию или отсутствию остей обусловлено одним геном. Безостость доминирует над остистостью.
- Признаки наследуются независимо, гены локализованы в разных хромосомах (или в одной, но на большом расстоянии).
- Генотипы исходных форм:
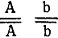 - плотный остистый колос,
- плотный остистый колос, 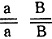 - рыхлый безостый колос, генотип
- рыхлый безостый колос, генотип 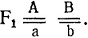
При скрещивании двух форм ячменя были получены результаты, представленные ниже:

Определить характер совместного наследования признаков; генотипы исходных форм и гибридов F1.
- Анализ по признакам
- Число рядов в колосе: в F1 единообразие, Р, очевидно, гомозиготны. В F2 расщепление: 485 - двурядные, 178 - четырехрядные. Н0 - расщепление по одному гену, 3:1, χ2 = 1,20, p>0,20. Отклонение случайно, гипотеза не отвергается. А - двурядный, а - четырехрядный колос.
- Наличие - отсутствие остей: в F1 единообразие, Р, очевидно, гомозиготный. В F2 расщепление: 501 - безостые, 162 - остистые. Н0 - расщепление по одному гену, 3:1, χ2 = 0,11, p>0,70. Отклонение случайно, гипотеза не отвергается. В - безостые, b - остистые колосья.
- Окраска чешуи: в F1 единообразие, Р, очевидно, гомозиготны. В F2 расщепление: 511 - черные, 152 - белые чешуи. Н0 - расщепление по одному гену, 3:1, χ2 = 1,52, p>0,20. Отклонение случайно, гипотеза не отвергается. С - черные, с - белые чешуи.
- Анализ совместного наследования признаков Поскольку по каждому признаку расщепление в F2 3:1 и в общем расщеплении 8 фенотипических классов без преобладания родительских комбинаций признаков, предполагаем (Н0) независимое наследование признаков с расщеплением - (3А:1а)×(3В:1b)×(3С:1с) = 27АВС:9АВс:9АbС:9аВС:3Аbс:3аВс:3abC:1abc. χ2 = 3,89, p>0,70. Отклонение случайно, гипотеза не отвергается.
- Различие исходных растений по количеству рядов в колосе обусловлено одним геном. Двурядность доминирует над четырехрядностью.
- Различие исходных растений по наличию остей обусловлено одним геном, безостость доминирует над остистостью.
- Различие исходных растений по окраске колоса обусловлено одним геном, черная окраска доминирует над белой.
- Признаки наследуются независимо, гены не сцеплены.
Генотипы исходных растении: 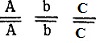 - двурядный остистый колос, черные чешуи,
- двурядный остистый колос, черные чешуи, 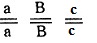 - четырехрядный безостый колос, белые чешуи; генотип растении
- четырехрядный безостый колос, белые чешуи; генотип растении 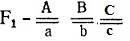
По теории вероятностей можно определить формулу расщеплений для двух, трех и более независимо наследуемых признаков. На основании этих формул легко вычисляется ожидаемая величина каждого класса в расщеплении (табл. VI.1).

Таблица VI.1. Формулы расщеплений при независимом наследовании двух признаков с моногенным контролем каждого из них
Очевидно, что отклонение в расщеплении по любому признаку, независимо от причин такого отклонения, будет влиять на характер совместного наследования признаков.
От скрещивания коротконогих кур, имевших простой гребень, с длинноногими петухами с гороховидным гребнем в F1 половина цыплят имела длинные, половина - короткие ноги, причем у всех были гороховидные гребни. В F2 от скрещивания потомков F1, различающихся по фенотипу, получили следующее расщепление: 55 с короткими ногами и гороховидным гребнем, 63 - с длинными ногами и гороховидным гребнем, 21 с короткими ногами и простым гребнем, 17 с длинными ногами и простым гребнем (∑ = 156). При скрещивании коротконогих потомков F1 между собой было также получено расщепление: 40 с короткими ногами и гороховидным гребнем, 23 с длинными ногами и гороховидным гребнем, 13 с короткими ногами и простым гребнем и 9 с длинными ногами и простым гребнем (∑ = 85). При скрещивании между собой длинноногих особей из F1 все потомки оказались длинноногими, из них 71 с гороховидным и 20 с простым гребнями. Объяснить расщепления, определить генотипы исходных форм и характер совместного наследования признаков.
1. Анализ по признакам
Поскольку в F1 произошло расщепление, можно утверждать, что один из родителей гетерозиготен; расщепление (примерно 1:1) свидетельствует о различии между исходными птицами по одному гену, χ2 = 0,1, p>0,70. Отклонение случайно, гипотеза не отвергается. Так как при скрещивании длинноногих особей расщепление отсутствует, можно предположить, что длинноногие птицы гомозиготны и это рецессивный признак (аа). Тогда коротконогость - доминантный признак, и коротконогие исходные птицы и потомки F1 имеют гетерозиготный генотип (Аа). Если это так, то расщепление в F2 от скрещивания гетерозиготных коротконогих кур из F1 53 коротко- и 32 длинноногих цыплят должно соответствовать менделевскому расщеплению 3:1, однако χ2 = 7,2, p<<0,01. Отклонение не случайно, гипотеза отвергается. Тот факт, что коротконогие при скрещивании между собой и с длинноногими всегда дают расщепление, позволяет предположить, что часть потомков, вероятно гомозиготных по аллелю А, погибает, и расщепление происходит по формуле 2:1; χ2 = 0,68, p>0,30. Отклонение случайно, гипотеза не отвергается. Генотип коротконогих кур Аа, длинноногих - аа.
б. Форма гребня: единообразие в F1 говорит о возможной гомозиготности исходных форм. Расщепление в F2: 252 гороховидных и 80 простых гребней позволяет предположить моногенный характер различий. χ2 по суммарным данным F2 от всех скрещиваний равен 0,15, p>0,50. Отклонение случайно, гипотеза не отвергается. В - гороховидный гребень, b - простой.
2. Анализ совместного наследования признаков
Из результатов анализа, проведенного по каждому признаку, можно записать генотипы исходных птиц: Aabb - коротконогие с простым гребнем, ааВВ - длинноногие с гороховидным гребнем. Потомки F1: AaBb - коротконогие с гороховидным гребнем и ааВb - длинноногие с гороховидным гребнем. Если гены наследуются независимо, то при скрещивании этих потомков между собой должно происходить расщепление, которое можно определить по теории вероятностей: (1А:1а)×(3В:1b) - 3АВ:3аВ::1Аb:1аb. χ2 = 0,99, p>0,80. Отклонение случайно, гипотеза не отвергается.
Скрещивание коротконогих потомков из F1 (AaBb×AaBb) должно привести к расщеплению: 6АВ:3аВ:2Аb:1ab - (2А:1a)×(3В:1b). χ2 = 0,91, p>0,80. Отклонение случайно, гипотеза не отвергается.
- Различие по длине ног между исходными формами обусловлено одним геном. Коротконогость обусловлена доминантным геном с рецессивным летальным действием, длинноногость - рецессивный признак.
- По форме гребня исходные особи также различаются по одному гену. Гороховидная форма гребня доминирует над простой.
- Признаки наследуются независимо. Гены локализованы в разных аутосомах (или в одной, но на большом расстоянии).
- Генотипы исходных форм:
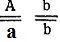 - коротконогие с простым гребнем,
- коротконогие с простым гребнем, 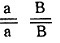 - длинноногие с гороховидным гребнем.
- длинноногие с гороховидным гребнем.
В скрещивании ржи с антоцианом и без антоциана было установлено, что признак контролируется одним геном с доминированием наличия антоциана (фиолетовая окраска проростков - Vi) над его отсутствием (зеленая окраска проростков - vi). Расщепление в F2 было: 986 фиолетовых и 301 зеленых проростков. χ2 = 1,77, p>0,10. В других скрещиваниях изучалось наследование красной и белой окраски ушков листа. Было обнаружено, что это различие обусловлено комплементарным взаимодействием двух независимо наследуемых генов (R1 и R2). Расщепление в опыте было: 720 с красными и 567 с белыми ушками. Н0 - расщепление 9:7, χ2 = 0,05, p>0,80.
Для выяснения характера совместного наследования признаков (наличие - отсутствие антоциана, красные-белые ушки) растения без антоциана с белыми ушками скрестили с растениями, имевшими антоциан и красные ушки. В F1 получили все растения с антоцианом и красными ушками, а в F2 расщепление:

Определить, как наследуются признаки. Как проверить правильность сделанных вами выводов?
- Анализ по признакам
- Наличие-отсутствие антоциана: в F1 единообразие, по-видимому исходные формы гомозиготны. В F2 расщепление: 425 с антоцианом и 147 без антоциана. Н0 - расщепление по одному гену, 3:1, χ2 = 0,15, p>0,50. Отклонение случайно, гипотеза не отвергается.
- Окраска ушков листа: в F1 единообразие - исходные формы по-видимому, гомозиготны. Расщепление в F2 - 248 с красными и 324 с белыми ушками- очевидно не согласуется с установленным ранее расщеплением (9 с красными : 7 с белыми ушками) по этому признаку. Можно предположить, что в изучаемом скрещивании проявляется новый тип взаимодействия между геном антоциана и генами окраски ушков (или в расщеплении участвует большее число взаимодействующих генов).
По теории вероятностей при независимом наследовании признаков можно рассчитать ожидаемое по ним расщепление в F2: (3/4 с антоцианом : 1/4 без антоциана)×(9/16 с красными : 7/16 с белыми ушками) = 27/64 с антоцианом и красными ушками: 21/64 с антоцианом и белыми ушками: 9/64 без антоциана с белыми ушками: 7/64 без антоциана с белыми ушками. В расщеплении, полученном в опыте, на достаточно большой выборке обнаружено только три фенотипических класса, отсутствует класс "без антоциана с красными ушками". Определив величину одного сочетания гамет -572:64=8,9, находим расщепление в опыте: 248:8,9 = 27,8; 177:8,9 = 19,9; 147:8,9 = 16,7, то есть примерно 27:21:16. χ2 = 0,92, p>0,50. Отклонение случайно, гипотеза не отвергается. Очевидно, что классы 9/64 и 7/64 имеют одинаковое фенотипическое проявление - без антоциана с белыми ушками листа. Рецессивный аллель гена, контролирующего антоциан (vi), подавляет проявление двух доминантных аллелей генов, контролирующих окраску ушков листа (R1 и R2). Красная окраска ушков проявляется при сочетании ViR1R2, все остальные сочетания трех генов обусловливают развитие белой окраски ушков листа:
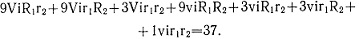
Расщепление в опыте -248 и 324 - соответствует ожидаемому 27 с красными: 37 с белыми ушками; χ2 = 0,32, p>0,50.
- Различие по окраске ушков листа между исходными формами обусловлено действием трех независимо наследуемых генов. Красная окраска проявляется только при наличии в генотипе доминантного аллеля гена, контролирующего наличие антоциана (Vi).
- Признаки наследуются независимо, гены не сцеплены.
- Генотипы исходных форм:
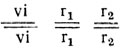 без антоциана с белыми ушками;
без антоциана с белыми ушками; 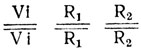 с антоцианом и красными ушками.
с антоцианом и красными ушками.
Хорошей проверкой правильности этих выводов послужило бы анализирующее скрещивание, для которого следовало использовать гетерозиготных по трем генам потомков F1, а в качестве анализатора- исходную форму без антоциана с белыми ушками. Нетрудно определить расщепление в таком скрещивании.
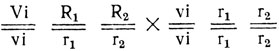

Следовательно, в Fa в данном случае должно быть 1/8 с антоцианом и красными ушками, 3/8 с антоцианом и белыми ушками, 4/8 без антоциана с белыми ушками.
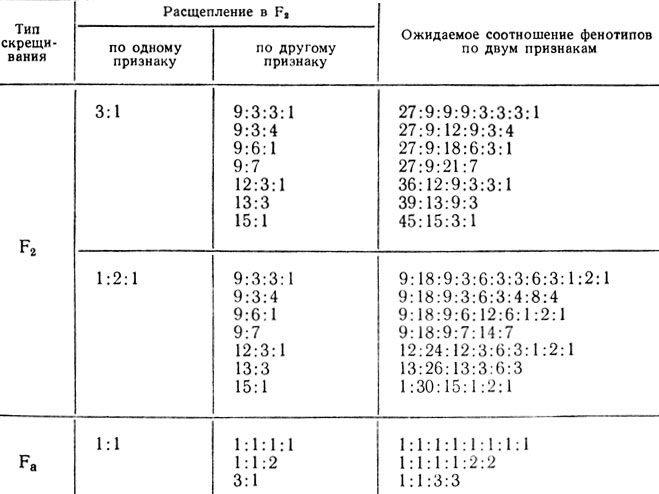
Таблица VI.2 Возможные расщепления в F2 и Fa при независимом наследовании двух признаков с разным генетическим контролем
Применение теории вероятностей позволяет находить формулы расщепления в случаях, когда признаки наследуются независимо и один (или каждый из них) контролируется двумя или большим числом взаимодействующих генов. В табл. VI.2 приведены возможные формулы расщеплений для двух признаков с разным генетическим контролем. Данные таблицы еще раз показывают, насколько усложняется анализ при участии в расщеплении нескольких генов, и как непросто увидеть характер расщепления без специальных приемов.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'