VII.1.5 Определение группы сцепления у растении на основе использования анеуплоидных серии - трисомиков, моносомиков и нуллисомиков
Анеуплоиды систематически появляются в скрещиваниях полиплоидов (2n×4n, 3n×3n и т. д.), при межвидовой гибридизации, ни могут возникать в результате нарушений в мейозе, например при нерасхождении хромосом. Это - трисомики (2n+1), моносомики (2n-1), тетрасомики (2n+1+1), нуллисомики (2n-2). У анеуплоидов избыточные или недостающие хромосомы принадлежат к одной гомологической паре. Диплоидные растения (2n) являются нормальными дисомиками, так как все хромосомы у них парные. Анеуплоидные серии используют для определения группы сцепления у растений.
а. Определение группы сцепления с помощью трисомиков
Трисомиков (2n+1) для генетического анализа обычно выделяют в потомстве триплоидных растений, где наблюдается большое разнообразие и наибольшая частота трисомиков по разным хромосомам, или их получают в скрещиваниях между диплоидными и тетраплоидными формами.
Трисомики у растений обычно менее плодовиты и жизнеспособны, чем дисомики. В мейозе у них образуются либо тривалент, либо бивалент и унивалент, либо три унивалента. Это зависит от вида растения и дополнительной хромосомы трисомика. Например, у кукурузы чаще встречаются триваленты, а у сахарной свеклы и томатов - бивалент и унивалент. Остальные хромосомы трисомика конъюгируют нормально, образуя биваленты. При распределении хромосом в мейозе у трисомиков образуются два типа гамет - n и n+1, причем дополнительная хромосома чаще передается через яйцеклетку, так как пыльцевые зерна с лишней хромосомой, как правило, не прорастают, либо их рост замедлен. Поэтому трисомиков используют в скрещиваниях в качестве материнских форм. Они могут иметь четыре разных генотипа по одному гену - AAA, AAa, Ааа, ааа. Расщепление по генам, локализованным в лишней хромосоме, происходит по тем же законам, что и у эуплоидов, т. е. имеет место как случайное хромосомное, так и случайное хроматидное расщепление с учетом двойной редукции (см. гл. V).
Для локализации гена с помощью этого метода последовательно скрещивают диплоидные формы, в генотипе которых содержится локализуемый ген, с набором трисомиков по всем хромосомам. В одном из этих скрещиваний, которое называется "критическим", появляется новый гетерозиготный трисомик. Для локализации рецессивной мутации диплоидную форму (аа) скрещивают с триплоидом, имеющим генотип AAA. Для локализации доминантной мутации используют трисомики с рецессивным генотипом (ааа). В первом поколении в любом из этих скрещиваний возникают новые трисомики с генотипами ААа или Ааа соответственно. Их выделяют либо по фенотипу, если они отличаются от диплоидов, либо цитологически. Затем самоопыляют или проводят анализирующее скрещивание с рецессивным диплоидом. Ниже приведены схемы скрещиваний для получения гетерозиготных трисомиков.
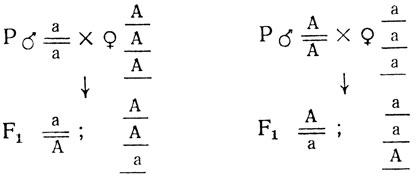
При случайном хромосомном расщеплении в потомстве от самоопыления трисомиков из F1 возникнут следующие расщепления:
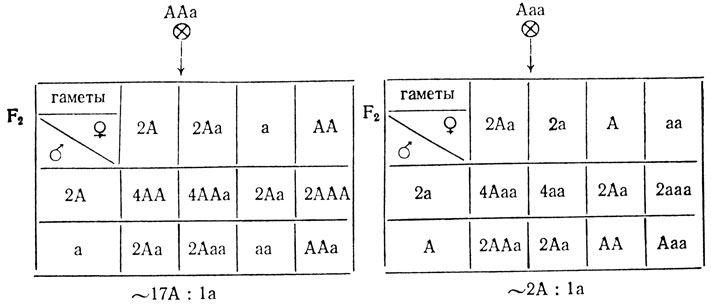
При случайном хроматидном расщеплении вероятность выщепления рецессива в F2 от самоопыления трисомика с генотипом ААа значительно меньше, так как он образует 3 типа гамет (С62) с разной вероятностью: 6АА, 1аа и 8Аа, что приведет к расщеплению 44А:1а.
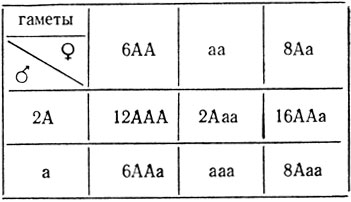
Аналогично можно рассчитать расщепление в Fa при случайном хромосомном и случайном хроматидном расщеплениях этих трисомиков.
Следует помнить, что результаты реальных скрещиваний редко совпадают с рассчитанными теоретически, так как у трисомика обычно образуется больше микро- и мегаспор с гаплоидным набором хромосом, а пыльца с набором n+1 часто стерильна или мало конкурентоспособна, обычно в потомстве трисомика преобладают дисомики и наблюдается дефицит рецессивов в критическом скрещивании.
Таким образом с помощью трисомиков удается локализовать гены в определенной группе сцепления, метод с успехом применяется на ряде диплоидных видов. Наиболее трудоемкая часть работы при его использовании - создание полной серии трисомиков.
б. Определение группы сцепления с помощью моносомиков и нуллисомиков
Моносомики - организмы с недостатком одной из хромосом (2n-1).
Поскольку у диплоидов моносомики, как правило, нежизнеспособны или мало жизнеспособны, моносомные формы используют для локализации генов лишь у аллополиплоидов, у которых гибридологический метод анализа очень сложен и с его помощью практически невозможно решить эту задачу.
На основе межвидовой гибридизации созданы серии моносомиков по всем хромосомам у аллотетраплоидного табака (Nicotiana tabacum) и у гексаплоидной мягкой пшеницы (Triticum aestivum), которая представляет собой сложный полиплоид (2n = 42). Первые моносомики мягкой пшеницы были получены Сирсом в 1939 г. от гаплоида, спонтанно возникшего среди гибридов F1 при скрещивании сорта пшеницы Чайниз Спринг (2n = 42) с рожью Secale cereale (2n = 14). У этого гаплоида было 28 хромосом (21 +7), и он имел стерильную пыльцу. После опыления его пыльцой пшеницы сорта Чайниз Спринг было получено тринадцать жизнеспособных семян. Среди растений, выросших из этих семян, пять оказались моносомиками, образующими в мейозе 20 бивалентов и один унивалент (2011+11). Среди остальных - шесть сказались дисомиками, нуллисомиками и моносомиками с различными аномалиями в мейозе. В дальнейшем Сирсом была создана полная серия моносомиков пшеницы сорта Чайниз Спринг, которая используется для получения новых серий моносомиков на других сортах. Для этого материнские моносомные растения сорта Чайниз Спринг скрещивают с отцовскими дисомными растениями из данного сорта. В потомстве от этих скрещиваний путем цитологического анализа отбирают моносомные растения, получившие свою единственную хромосому от отца. Эти растения вновь скрещивают с отцовским сортом и такой бэккросс повторяют не менее шести раз, поэтому новые моносомики становятся идентичными отцовскому сорту.
Для определения группы сцепления последовательно скрещивают диплоидные растения, гомозиготные по локализуемому гену, со всеми моносомными линиями сорта. В потомстве от этих скрещиваний цитологически идентифицируют вновь возникших моносомиков. Если рецессивная мутация полностью проявляется в гемизиготном состоянии, то все моносомики F1 в скрещивании 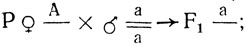
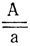 будут иметь мутантный фенотип и группа сцепления определяется уже в первом поколении. Иногда рецессивный фенотип не проявляется в гемизиготе и моносомные потомки такого скрещивания (а) имеют нормальный фенотип. Моносомики, возникающие в F1 при скрещивании
будут иметь мутантный фенотип и группа сцепления определяется уже в первом поколении. Иногда рецессивный фенотип не проявляется в гемизиготе и моносомные потомки такого скрещивания (а) имеют нормальный фенотип. Моносомики, возникающие в F1 при скрещивании 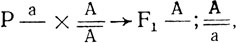 неотличимы от дисомиков и также имеют нормальный фенотип. В этих случаях локализацию генов проводят на основе расщепления в F2, получаемого от самоопыления моносомиков F1, выявляемых цитологически. В потомстве моносомика _а рецессивный фенотип может проявиться либо у гомозиготных дисомиков
неотличимы от дисомиков и также имеют нормальный фенотип. В этих случаях локализацию генов проводят на основе расщепления в F2, получаемого от самоопыления моносомиков F1, выявляемых цитологически. В потомстве моносомика _а рецессивный фенотип может проявиться либо у гомозиготных дисомиков 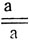 , либо у нуллисомиков, лишенных этой хромосомы. В потомстве моносомика А рецессивный фенотип проявляется только у нуллисомиков. Моносомики, дисомики и нуллисомики выщепляются в потомстве самоопыленных моносомных растений, образующих два сорта гамет - n и n-1. Соотношение этих типов гамет и соответственно, образование моно-, ди- и нуллисомных растений зависит, главным образом, от сорта. Так, через материнские моносомные растения сорта Чайниз Спринг передается от 70 до 81% гамет с недостающей хромосомой (в среднем - 75%), а через отцовские - лишь 4%, благодаря чему в F2 возникает следующее соотношение разных типов растений.
, либо у нуллисомиков, лишенных этой хромосомы. В потомстве моносомика А рецессивный фенотип проявляется только у нуллисомиков. Моносомики, дисомики и нуллисомики выщепляются в потомстве самоопыленных моносомных растений, образующих два сорта гамет - n и n-1. Соотношение этих типов гамет и соответственно, образование моно-, ди- и нуллисомных растений зависит, главным образом, от сорта. Так, через материнские моносомные растения сорта Чайниз Спринг передается от 70 до 81% гамет с недостающей хромосомой (в среднем - 75%), а через отцовские - лишь 4%, благодаря чему в F2 возникает следующее соотношение разных типов растений.

Если моносомик имеет генотип а, то в F2 возникнет соотношение: 73% растений с нормальным фенотипом (моносомики) и 27% с рецессивным фенотипом (дисомики и нуллисомики). Моносомик с генотипом А в F2 будет давать иное соотношение: 97% нормальных растений (дисомики и моносомики А) и 3% -рецессивных (нуллисомики).
Эти частоты приняты в качестве стандарта при оценке расщеплений в потомстве самоопыленных моносомиков.
Если ген локализован не в моносомичной хромосоме, то расщепление происходит как обычно - 3:1 или 1:2:1.
При использовании серий нуллисомиков для определения группы сцепления скрещивают дисомные растения, гомозиготные по локализуемому гену, с нуллисомными линиями данного сорта. Все потомство от таких скрещиваний оказывается моносомическим по какой-либо хромосоме. Если моносомические потомки, полученные от скрещивания рецессивной формы с нуллисомиком, окажутся рецессивными по этому признаку, значит ген локализован в хромосоме, отсутствующей у нуллисомика.
Локализация рецессивных мутаций по некоторым признакам может быть проведена непосредственно по фенотипу нуллисомика. Например, нуллисомик по 16-й хромосоме у краснозерного сорта пшеницы дает белые зерна. Следовательно, аллель, контролирующий красную окраску, локализован в 16-й хромосоме.
С помощью серий анеуплоидов к настоящему времени локализованы гены, контролирующие содержание белка, особенности пигментации зерна, устойчивости к наиболее вредным заболеваниям, яровость - озимость и другие важные признаки пшеницы. Использование этого метода ограничено - он пригоден только для аллоплоидных видов.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'