X.3.3. Анализ транслокаций
Транслокации - аберрации обменного типа, в них участвуют две или больше хромосом. Асимметричные воссоединения образующихся фрагментов - нереципрокные транслокации - приводят к гибели клеток и проявляются как доминантные летали.
При симметричных обменах (реципрокные транслокации) образуются новые группы сцепления, в каждой хромосоме сохраняется по одной центромере. Как правило, реципрокные транслокации фенотипически не проявляются ни в гомо-, ни в гетерозиготном состоянии, за исключением случаев, когда они вызывают эффект положения. Их можно обнаруживать с помощью цитологического и гибридологического методов.
Цитологически транслокации выявляются в мейозе у гетерозигот по наличию характерных фигур (креста в момент конъюгации хромосом, а также колец и восьмерок при расхождении гомологов) (см. гл. VII). Как уже указывалось, у растений, гетерозиготных по транслокации, может наблюдаться частичная стерильность за счет гибели пыльцы с несбалансированными хромосомами, возникающими при расхождении хромосом из кольца. У разных растений степень стерильности при этом различна. Она зависит от частоты формирования колец в метафазе первого деления мейоза. Например, у кукурузы стерильность пыльцы у гетерозигот по транслокации составляет примерно 50%, т. е. она проявляется как доминантный признак. У пшеницы, дурмана, томатов и ряда других растений чаще происходит ориентация хромосом в метафазе I в виде восьмерок, поэтому гетерозиготы по транслокации у них более фертильны.
Транслокации встречаются и у животных, например у кузнечиков, скорпионов, дрозофилы и др. Однако у животных гаметы с несбалансированными хромосомами - это зиготические летали. При отсутствии кроссинговера у особей одного из полов (самцы дрозофилы, самки шелкопряда) транслокации всегда сохраняются и могут быть выявлены в скрещиваниях.
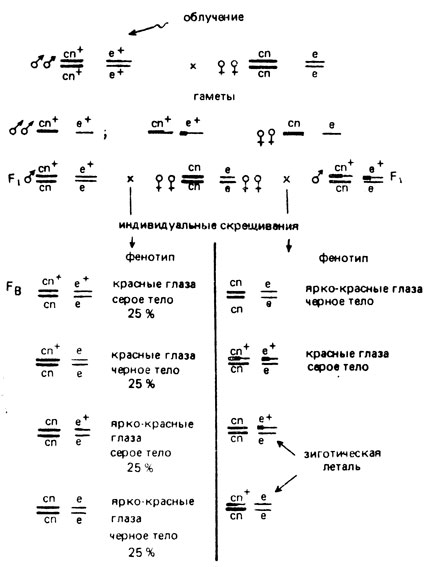
Рис. X.9. Схема эксперимента, проводимого для обнаружения транслокаций у дрозофилы
У дрозофилы для обнаружения транслокаций, возникающих в сперматозоидах после обработки мутагеном самцов дикого типа, их скрещивают с самками из линии-анализатора, содержащей маркеры по разным хромосомам (например, по второй хромосоме cn, и е - по третьей). Каждого гетерозиготного самца F1, полученного в этом скрещивании, возвратно скрещивают с 1-2 самками из линии-анализатора. Все потомки такого возвратного скрещивания фенотипически нормальны. Если гетерозиготный самец F1 не имеет в своем генотипе транслокации, то в Fb возникает четыре типа потомков в равном количестве. Если самец F1 оказывается гетерозиготным по транслокации, то в Fb обнаруживается только два класса в расщеплении - нормальные и рецессивные (cn, e) мухи: гены исследуемых хромосом как бы проявляют полное сцепление (рис. X.9). Проведя учет культур, в которых появились только два класса мух, и отнеся их к общему числу проанализированных культур, определяют частоту (%) возникновения транслокаций между 2- и 3-й хромосомами дрозофилы.
Если транслокация затрагивает небольшие участки хромосом, то иногда могут выживать зиготы с несбалансированным набором хромосом. При этом гены, которые у исходных форм наследовались сцепленно, проявят независимое наследование, а гены, ранее наследовавшиеся независимо, окажутся сцепленными.
У самок дрозофилы, гетерозиготных по транслокации, может проходить кроссинговер с участием транслоцированных хромосом, однако он заметно подавлен вследствие гибели части кроссоверных гамет. Часто транслокации служат причиной различий в числе хромосом у близких видов. Они основа для получения сбалансированных леталей, в которых поддерживается постоянная гетерозиготность (например, у энотеры).
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'