X.3.2. Анализ инверсий
Инверсии - результат двух разрывов в одной хромосоме и воссоединения их в новом положении вследствие поворота участка между разрывами на 180°. Перицентрические инверсии изменяют положение центромеры, парацентрические ее не затрагивают. Инверсии могут полностью или частично перекрывать друг друга или возникают независимо в разных участках хромосомы. Хотя инверсии не нарушают баланса генов, они изменяют порядок расположения генов в хромосоме, что может вызывать эффект положения, т. е. изменять регуляцию действия генов, локализованных рядом с точками разрывов. Инверсии выявляются с помощью цитологических методов, а также при гибридологическом анализе.
Цитологическими методами инверсии обнаруживаются на митотических хромосомах благодаря дифференциальной окраске по изменению расположения морфологических маркеров - центромер, хромомер и пр.; инверсии приводят к образованию мостов и фрагментов в анафазе первого мейотического деления; появление инвертированных петель на политенных хромосомах слюнных желез у насекомых также свидетельствует об их возникновении.
Инверсии являются "запирателями" кроссинговера - в потомстве гетерозигот по инверсии резко снижается процент рекомбинантных особей по генам, локализованным в инвертированном участке. Почему это происходит? Если инверсия затрагивает небольшой участок, то на нем отсутствует кроссинговер. Если инвертирован большой участок, конъюгация осуществляется с образованием инвертированной петли, внутри которой может происходить кроссинговер. Однако зиготы, образующиеся с участием гамет, в которые попадают продукты кроссинговера, оказываются нежизнеспособными, так как эти гаметы несут несбалансированные хромосомы: у гетерозигот по инверсии в результате одиночного кроссинговера в парацентрической инверсии в анафазе первого деления мейоза возникают дицентрик и акроцентрик; во втором мейотическом делении дицентрик разрывается, а акроцентрик обычно лизируется (рис. X.7, а). При одиночном кроссинговере в перицентрической инверсии также образуются гаметы с делециями и дупликациями (рис. Х.7, б). Жизнеспособные рекомбинанты в потомстве гетерозигот по инверсии возникают только в том случае, когда внутри инверсии происходит двойной кроссинговер (рис. Х.7, в).
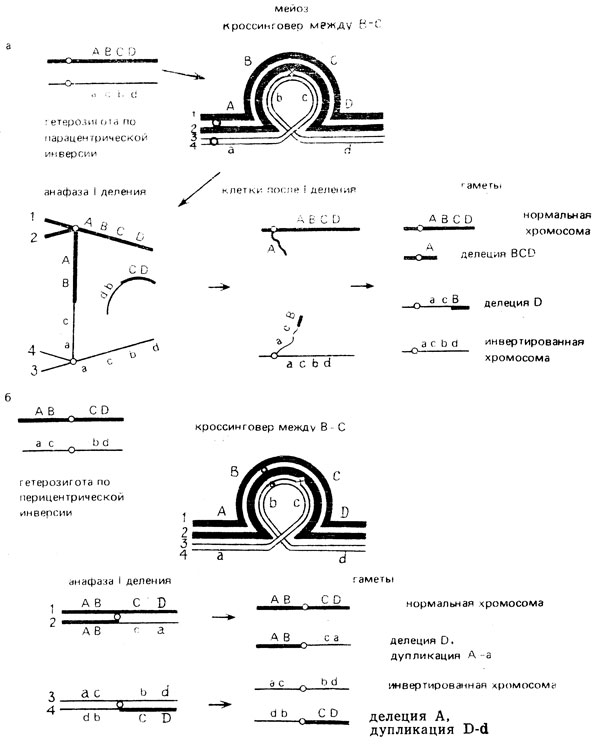
Рис. X.7. Кроссинговер в парацентрической (а) и перицентрической (б) инверсиях. Образование мостов, делений и дупликаций в анафазе 1 мейоза
У организмов, гетерозиготных по инверсии, как правило, снижена плодовитость. Показателем инверсии в гетерозиготе у какой-либо особи служит резкое снижение рекомбинантных потомков, подавление кроссинговера. Сопоставляя частоту рекомбинации в опыте по генам с известной локализаций с расстояниями между этими генами по стандартной генетической карте, выявляют инверсии и локализуют точки их разрывов. Рассмотрим конкретный пример такого анализа.
Гомозиготных самок, маркированных пятью рецессивными мутациями, локализованными во второй хромосоме, скрестили с самцами дикого типа, облученными рентгеновскими лучами. Каждую гетерозиготную самку из F1 возвратно скрестили с гомозиготными рецессивными по исследуемым генам самцами (Fa):
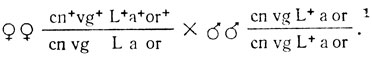
1 (cn - cinnabar - ярко-красные глаза, 57,5; vg - vestigial - зачаточные крылья, 67; L - Lobe - лопастные глаза, 72; а - arc - широкие изогнутые крылья, 99,2; or - orange - оранжевые глаза, 106,7. Все гены локализованы в правом плече хромосомы 2.)
Опыт поставлен для выявления инверсий, которые могли возникнуть в сперматозоидах облученных самцов. На основании анализа расщеплений в потомстве нескольких самок обнаружили резкое снижение частоты кроссинговера между некоторыми генами. Результаты опыта и расстояния между исследуемыми генами на стандартной генетической карте приведены ниже:

Определить, есть ли инверсия в генотипе исходных самок; если есть, то какие локусы второй хромосомы затронуты инверсией.
Частоты кроссиногвера между генами cn и vg, а и or довольно точно совпадают с расстояниями на стандартной генетической карте. Частоты кроссинговера между генами vg-L и L-а значительно ниже, чем на стандартной карте. Можно предположить, что в генотипе исследованных гетерозиготных самок имеется инверсия, включающая гены vg, L и а.
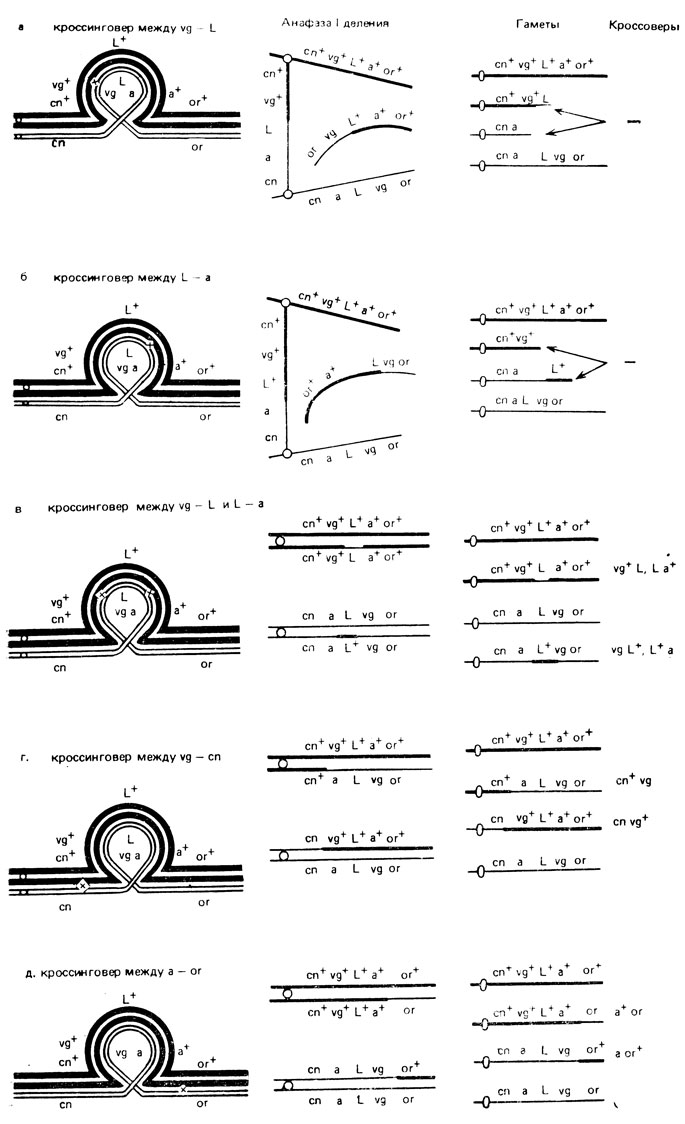
Рис. X. 8. Образование кроссоверов у гетерозигот по инверсии по генам, локализованным в инвертированном участке. Стрелками указаны хромосомы, не попадающие в гаметы (а, б, в, г, д)
Одиночный кроссинговер в инверсии между генами vg и L должен привести к образованию дицентрика и акроцентрика и, следовательно, к формированию рекомбинантных гамет с несбалансированными хромосомами. Аналогичный результат получается при кроссинговере между генами L и а (рис. X.8, а, б). Кроссоверы по этим парам генов появляются только в случае двойного кроссинговера между генами vg и L и L-а внутри инверсии (рис. Х.8, в). В участках за пределами инверсии, где локализованы гены cn и or, кроссинговер проходит нормально, и образуются жизнеспособные кроссоверные гаметы (рис. Х.8, г, д).
Снижение частоты кроссинговера между генами vg - L и L-a связано, по-видимому, с парацентрической инверсией, затрагивающей локусы vg, L и а. Для окончательного вывода о наличии такой инверсии следует провести цитологический анализ хромосом слюнных желез гетерозиготных самок.
Как уже указывалось, инверсии как "запиратели" перекреста вводятся в генотип линий-анализаторов, применяемых для выявления вновь возникших мутаций и определения групп сцепления и др.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'