X.3.1. Анализ делеций и дупликаций
Нехватки концевых участков хромосом и делеции (выпадение и утрата внутренних участков хромосом), возникающие в генеративных клетках, в гомо- и гемизиготном состоянии, а также у гаплоидов, как правило, летальны, но могут сохраняться в гетерозиготе.
Гетерозиготы по нехваткам и делециям обладают плейотропным действием, снижают плодовитость и жизнеспособность особей. Если делеции затрагивают генетически активные районы, они почти всегда проявляются фенотипически, как, например, упоминавшаяся в гл. VIII мутация Notch у дрозофилы, на которой выявлено более 130 делеции разной протяженности типа Notch. Установлено, что разные аллели в гемизиготном состоянии у самок (делеционные мутанты) могут проявляться не так, как у нормальных гемизиготных самцов.
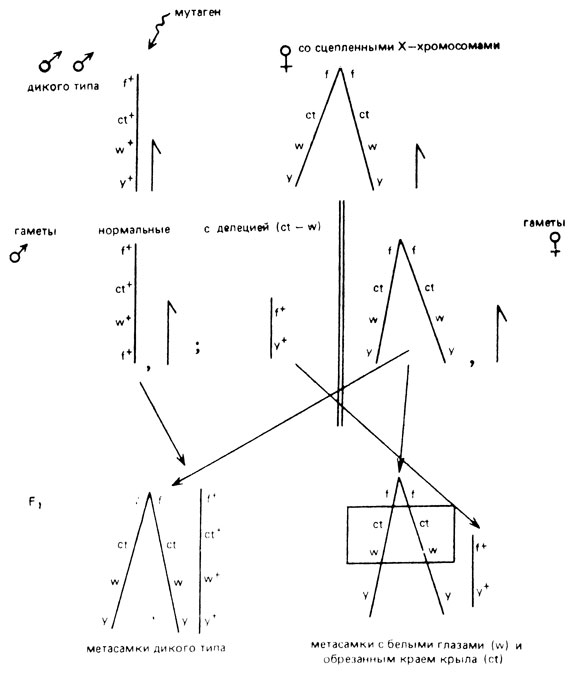
Рис. Х.6. Выявление крупных делений в Х-хромосоме дрозофилы с помощью сцепленных Х-хромосом. Рецессивные мутации: f(forked) - вильчатые щетинки, ct(cut) - обрезанный край крыла, w(white) - белые глаза, y (yellow) - желтое тело
Крупные делеции в Х-хромосоме дрозофилы выявляют методом сцепленных Х-хромосом. Для этого самки 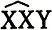 маркируются несколькими рецессивными мутациями и скрещиваются с мутагенизированными самцами дикого типа. В их потомстве могут возникать суперсамки (метасамки)
маркируются несколькими рецессивными мутациями и скрещиваются с мутагенизированными самцами дикого типа. В их потомстве могут возникать суперсамки (метасамки) 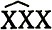 , которые при отсутствии делеции в Х-хромосоме, полученной от самца, должны быть дикого типа. Если у самца образуются делеция, захватывающая область маркеров, то у самок проявляется один или несколько рецессивных маркеров (рис. Х.6).
, которые при отсутствии делеции в Х-хромосоме, полученной от самца, должны быть дикого типа. Если у самца образуются делеция, захватывающая область маркеров, то у самок проявляется один или несколько рецессивных маркеров (рис. Х.6).
Прекрасным объектом для обнаружения делеций служат гигантские политенные хромосомы некоторых насекомых - Drosophila, Chironomus и др. (см. гл. VIII, рис. VIII.2). Своеобразный метод учета делеций на участке, разделяющем два адениновых гена (ade 3A и ade 3B), предложен на нейроспоре. Метод основан на том, что мутанты по этим генам растут на среде с аденином только при наличии этого участка. Если он делетирован, то мутации не растут даже на среде, обогащенной аденином. Критерием, с помощью которого выявляют эти делеций, является отсутствие роста адениновых мутантов на среде с аденином. Делеций генов ade ЗА и ade 3B этим методом не выявляются.
Для выявления делеций на некоторых растениях, например на кукурузе, применяют так называемый "тест на специфические локусы" - аллели, контролирующие рецессивные признаки эндосперма: окраску, форму, наличие или отсутствие крахмала и пр. Тест основан на выявлении эффекта "псевдодоминирования" у гетерозигот по этим маркерам. Для этого подвергают мутагенной обработке пыльцу нормальных растений и опыляют ею женские растения, маркированные несколькими рецессивными мутациями по признакам эндосперма. О наличии делеций узнают по проявлению рецессивных признаков на гибридных семенах М1. Очевидно, что можно проанализировать большое число зерен в початках. Однако этот тест не позволяет количественно оценить частоту возникновения делеций в спермиях, так как в зрелом пыльцевом зерне у ряда растений генеративное ядро уже разделено на два ядра и генотип второго ядра, сливающегося с яйцеклеткой, остается неизвестным. Если же обрабатывать мутагеном клетки микроспор, то можно определить частоту их возникновения, так как в микроспоре оба ядра спермиев имеют одинаковый генотип. Поэтому эндосперм и зародыш оказываются гетерозиготными по одной и той же делеций. Кроме того, у растений, гетерозиготных по делеций, происходит гибель части гамет, и растения M1 образуют до 50% абортивной пыльцы.
Эффект "псевдодоминирования" используют в некоторых случаях для выявления делеций у мышей.
Наличие делеций у диплоидных организмов выявляют также по отсутствию рекомбинаций между сцепленными маркерами, она приводит к уменьшению расстояния на генетической карте между крайними генами.
К сожалению, далеко не всегда удается отличить делецию от генной мутации. Четкий критерий делеций - отсутствие обратного мутирования и рекомбинирования с определенными мутациями.
В результате разрывов хромосом, неравного кроссинговера, а также благодаря избыточной репликации участков хромосом и последующего их включения в хромосому возникают дупликации. Они бывают внутрихромосомными (прямые, инвертированные, смежные, тандемные) или межхромосомными. Гены гистонов, тРНК, рРНК и др., повторяющиеся в геноме эукариот сотни раз и относящиеся к классу умеренных повторов, представляют собой дупликации, которые достаточно легко обнаруживаются на политенных хромосомах (см. гл. VIII).
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'