§ 4. Ген с позиций молекулярной биологии
Ген - элементарный, дискретный, материальный наследственный фактор, который определяет строение одной белковой полипептидной цепи. Он является единицей наследственности и передается от родителей к их потомкам.
Классическая генетика называла геном отдельный участок (локус) хромосомы, ответственный за реализацию определенного наследственного признака. В начале 20-го века генетики представляли, что гены неизвестной химической природы располагаются линейно вдоль хромосомы (см. главу IV, § 5), подобно "бусинкам", нанизанным на нить. В наши дни доказано, что ген представляет собой определенную последовательность нуклеотидов в нуклеотидной цепи ДНК. Гены, так же как и триплеты, в молекуле ДНК расположены линейно. В последнее время специальными приемами удалось изолировать некоторые гены из бактерий в чистом виде. При помощи электронного микроскопа удалось также сфотографировать процесс транскрипции, демонстрирующий работу определенных генов, переписывающих свою нуклеотидную последовательность на комплементарную последовательность РНК.
Представляя собой участок ДНК, каждый ген состоит из некоторого числа мономеров - нуклеотидов. Размеры генов неодинаковы. Они зависят от размера белка, который кодирует данный ген: если белок состоит, например, из 200 остатков аминокислот, то в гене будет 600 пар нуклеотидов, что соответствует относительной молекулярной массе около 420 000.
Согласно современным представлениям, каждая хромосома содержит одну гигантскую молекулу ДНК. Такая молекула ДНК - генофора ("хромосома") - бактерии кишечной палочки замкнута в кольцо окружностью 1,2 мм, содержащее около 4*106 нуклеотидных пар. В хромосоме млекопитающих и человека средняя длина ДНК составляет около 2 см, что соответствует около 5,3*107 пар нуклеотидов. Всего же в геноме человека (в каждой клетке) около 6*10-12 г ДНК, что соответствует общей длине около 2 м (5,3*109 пар нуклеотидов) и молекулярной массе около 3,7*1012 (табл. 2).

Таблица 2. Некоторые характеристики молекул ДНК различных организмов
Учитывая, что средняя длина структурного гена составляет 500-1000 нуклеотидов, ДНК человека теоретически могла бы кодировать около 10 млн. белков.
В действительности за время существования генетики описано множество генов. Так, у хорошо изученной плодовой мушки (Drosophila melanogaster) их описано 3-4 тыс., у человека по приблизительным подсчетам число генов составляет от 10 тыс. до 100 тыс. Таким образом, в нашем геноме имеется огромный "избыток" ДНК.
В настоящее время показано, что значительная часть этой "избыточной" ДНК содержит гены, не являющиеся структурными, т. е. не кодирующие структуру определенных белков, а выполняющие целый ряд функций, связанных с регуляцией активности тех или иных участков генома. Помимо этого, многие, в особенности регуляторные, гены многократно повторяются, т. е. геном содержит много (иногда десятки тысяч) копий одного и того же гена. Наконец, довольно большие участки ДНК разделяют гены между собой, кодируют начало и конец транскрипции и выполняют ряд других, еще не полностью выясненных функций. Кроме этого, следует помнить, что у многоклеточных, в частности у человека и млекопитающих, в результате сложного процесса дифференцировки в ходе индивидуального развития образуется много разнообразных органов и тканей. Состоят эти органы и ткани из многочисленных клеток, специализированных для выполнения соответствующих функций. Видимо, именно поэтому большая часть генов многоклеточных организмов находится в неактивном состоянии.
Каково же тонкое строение гена? Согласно определению классической генетики, ген - это структурная единица наследственности, определяющая данный признак (теперь - полипептидную цепь), способная рекомбинироваться и мутировать. Американский исследователь С. Бензер исследовал тонкое строение гена на участке rII бактериофага Т4 кишечной палочки. Ему удалось получить много однозначных мутаций, дававших совершенно одинаковое изменение фага, которые, однако, располагались в разных участках одного и того же гена. Подобно этому, один и тот же ген давал много рекомбинаций, локализованных в разных участках. Размер каждого участка, который дает мутацию (см. главу V) или обладает способностью к комбинациями с другим участком (рекомбинациям), оказался намного меньше, чем размер гена, определяющего структуру полипептидной цепи. В результате этой и других схожих работ единое понятие ген приходится расчленить на три более частных понятия: ген, определяющий структуру полипептидной цепи,- цистрон, или структурный ген; ген, способный мутировать,- ген мутации, или мутон; ген, способный к рекомбинациям,- ген рекомбинации, или рекон.
Если размер цистрона составляет в среднем 500-1000 пар нуклеотидов, то размер рекона, а также мутона не превышает одной - двух пар нуклеотидов, поскольку изменение даже одного основания в ДНК способно вызвать так называемую точковую мутацию.
Изучая индукцию ферментов у бактерии, например индукцию фермента β-галактозидазы, после добавления в среду лактозы французские ученые Ф. Жакоб и Ж. Моно получили ряд мутантов с нарушенной регуляцией работы генов. Среди этих мутантов были такие, которые вообще не синтезировали β-галактозидазу, но были и такие, которые синтезировали ее независимо от присутствия индуктора, т. е. такие, у которых вследствие мутации был нарушен механизм индукции. При таких нарушениях соответственно был нарушен синтез других ферментов, связанных с превращением лактозы.
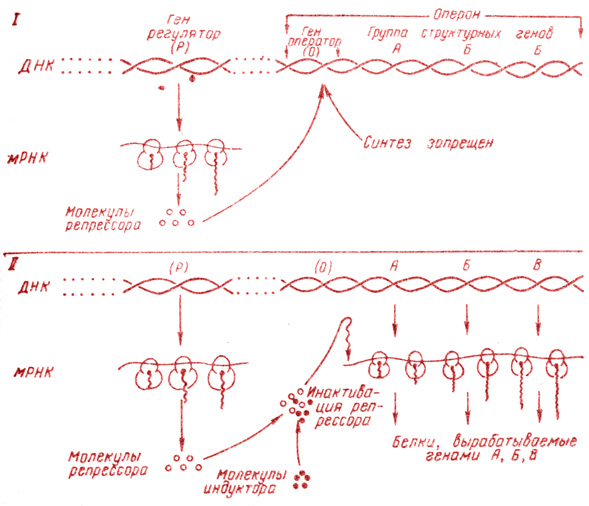
Рис. 5. Схема регуляции синтеза ферментов (белков) путем отрицательной индукции. Два состояния оперона: I - репрессированное; репрессор занимает место на опероне (О), прекращая этим синтез мРНК; II - индуктор приводит репрессор в неактивное состояние, структурные гены начинают функционировать
На основании своих исследований Жакоб и Моно предложили схему регуляции синтеза ферментов (белков) у микроорганизмов путем так называемой отрицательной индукции (рис. 5). Согласно этой схеме, ряд структурных генов (цистронов), кодирующих синтез ферментов, сопряженных между собой в одном и том же метаболическом процессе, расположены последовательно друг за другом в одной полинуклеотидной цепи. Эти структурные гены вместе с особым геном - оператором соединены в общую единицу - оперон. Помимо оперона, в том же или другом месте генома имеется особый ген - регулятор. Этот ген кодирует образование особого агента - репрессора, соединяющегося с оператором и подавляющего функцию всего оперона. Низкомолекулярное вещество индуктор (в случае лактозного или lac-оперона это лактоза или какой-либо другой β-галактозид) соединяется с репрессором и инактивирует его, вследствие чего последний теряет способность подавлять функцию оперона. В настоящее время lac-репрессор выделен в чистом виде и оказался белком. (Кроме оператора, в составе оперона обнаружен еще один участок, необходимый для начала транскрипции,- промотор.).
Способы генетического контроля биосинтеза белков могут быть более сложными. В некоторых случаях сами ферменты могли играть роль репрессора, не все структурные гены ферментов данного метаболического пути оказались соединенными в опероны.
У высших организмов генетический контроль и регуляция биосинтеза белков много сложнее, чем у микроорганизмов. Прежде всего, хотя геном одинаков в разных клетках и тканях организма, в них синтезируются разные белки (например, миозин и другие белки - в мышцах, пепсин - в слизистой оболочке желудка, гемоглобин - в эритробластах и ретикулоцитах и т. д.). Это явление объясняется блокированием большей части генов в процессе дифференцировки, осуществляемым, по-видимому, при участии гистонов и других белков хроматина. Помимо этого, имеет место также регуляция, связанная с индукцией ряда ферментов и других белков. Эта индукция у высших часто осуществляется на уровне всего организма при помощи гормонов, нервных импульсов и других механизмов.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'