Невидимые хромосомы
Что скрывается за термином "генетический материал" микроба? В каком направлении идет обмен этим материалом - от круглой клетки к овальной или наоборот? Какая клетка отцовская, а какая материнская? Где записана генетическая информация у микробов? Есть ли у них хромосомы, и, если есть, как они устроены? (фото 7).
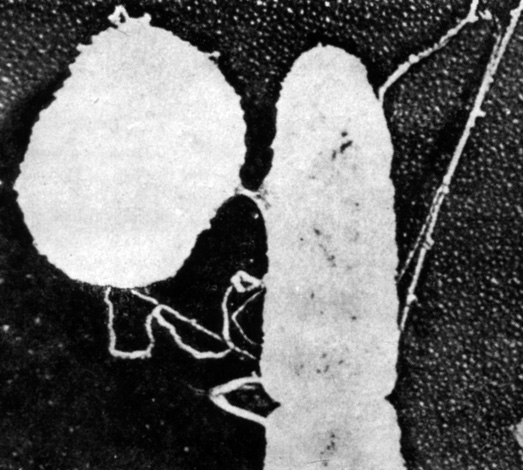
Фото 7. Момент конъюгации двух клеток кишечной палочки К12. Слева - бактерия-мать, справа - Hfr. Виден мостик, по которому следует хромосома
Все эти вопросы столпились вокруг первых картографов микробных хромосом. И начали они с последнего...
Хромосома бактерии... В начале 50-х годов это звучало как гром среди ясного неба. Ведь еще даже не было решено, есть ли у микробов ядро, а о хромосомах и говорить нечего. И все-таки Ледерберг решил попытаться... Он чувствовал, что тайна клеточного ядра бактерий может раскрыться вместе с тайной микробных гибридов, и не побоялся пуститься в путь.
Вход в лабиринт не таил никаких опасностей. Ледерберг взял в проводники уже знакомых нам "разведчиков" - мутантов кишечной палочки. Но они отличались друг от друга не только по способности к синтезу пяти важных метаболитов, но еще и по чувствительности к фагу Т1 - вирусу, способному растворять некоторые культуры кишечных палочек.
Удачливому экспериментатору посчастливилось получить четырех мутантов. Давайте для наглядности нарисуем их. Темным обозначим признаки, которых нет у искалеченных мутацией бактерий.
Внимание! Теперь самый ответственный момент. Скрещивается первая пара. То, что получатся полноценные, способные синтезировать "все что угодно" гибриды, не вызывало сомнений. Но будут ли они защищены от фага? Этого никто не мог сказать заранее. В результате опыта две трети полноценных потомков-рекомбинантов оказались устойчивы к фагу, и лишь одна треть - чувствительна.
А вот при скрещивании второй пары отношение к фагу стало иным: устойчивой была одна треть рекомбинантов, а чувствительными - почти две трети. В чем же дело? Ведь пары были одинаковы во всем. Во всем, за единственным, на вид несущественным, исключением: "фаговый", если можно так сказать, признак несли разные мутанты. И здесь Ледерберг увидел краешек истины: признак чувствительности или устойчивости к фагу "привязан" к признакам синтеза треонина (T), лейцина (L) и витамина B1! Рекомбинант наследует этот признак вместе с названными тремя. Иначе не могли бы 2/3 потомков первого скрещивания оказаться устойчивыми к фагу Т1. Признаки бактерий сцеплены друг с другом в определенном, строгом порядке. Скорее всего, как на линейке. Сидящие ближе друг к другу передаются вместе, далеко расположенные - порознь.
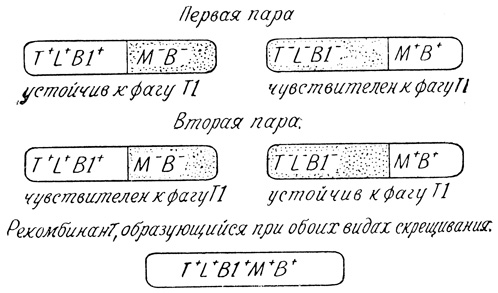
Схема опыта Ледерберга
Следовательно, если у бактерий признаки расположены линейно, то они имеют хромосому. И можно построить... ее карту.
Опыты, споры, неимоверный труд по изучению сотен мутантов. Шаг за шагом изучал ученый таинственную карту хромосомы бактерий. И вот перед его взором с тонкой линии - карты хромосомы бактерий - стали исчезать белые пятна. На нее ложились все новые и новые гены.
Гибриды Ледерберга принесли в микробиологию неслыханные дотоле вести. У бактерий тоже есть хромосомы, но не два набора, а всего один. Если помните, в обычных, неполовых клетках высших организмов каждая хромосома имеет свою пару - близнеца" И ученый искал такого близнеца бактериальной хромосомы. Это была на редкость трудная задача: ведь цитологу найти вторую хромосому ничего не стоит, ее можно просто увидеть под микроскопом, а бактериальную хромосому, как известно, увидеть невозможно. Но в стране генетических рекомбинаций можно наблюдать и невидимое. Ледерберг и Татум предположили, а затем доказали: хромосома у бактерий одна, ранняя стадия (диплоидная) у оплодотворенных клеток недолговечна.
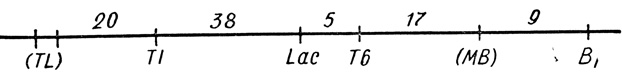
Первая генетическая карта кишечной палочки, составленная Ледербергом. Числа указывают относительную частоту рекомбинаций между отдельными признаками
Эти опыты полностью соответствовали наблюдениям американской исследовательницы Эвелины Виткин.
Она изучала мутации, возникающие у кишечной палочки при облучении ультрафиолетом. Как известно, в норме кишечная палочка сбраживает лактозу, а под влиянием облучения можно получить бактерии, лишенные этого качества. На специальной среде с индикатором нормальные микробы образуют ярко-красные колонии, а мутантные - бесцветные.
И вот что удивило исследовательницу: если облучать культуру в начальной стадии роста, то появятся несколько пестрых колоний, наполовину красных, наполовину бесцветных. Если же облучать культуру в стадии активного роста (так называемая лог-фаза), то можно увидеть колонии, окрашенные уже на три четверти.
Виткин знала, что покоящаяся культура содержит клетки с одним нуклеоидом, культура, начинающая делиться, - с двумя, культура в лог-фазе - с четырьмя. И вот что любопытнее всего: отношение числа клеток, содержащих один нуклеоид, к числу клеток с двумя нуклеоидами было пропорционально отношению количества окрашенных колоний к количеству двухцветных колоний. Та же закономерность выявилась и для колоний, окрашенных на три четверти.
Виткин рассуждала примерно так. В бактерии с двумя нуклеоидами мутация может поразить только один, и в нем нарушится работа гена по синтезу фермента, сбраживающего лактозу. Но ведь второй-то нуклеоид остается цел! Когда клетка разделится, то одной из дочерей достанется мутантный нуклеоид (и его потомство даст бесцветную колонию), другой- нормальный (и его потомство даст красную колонию). Впрочем, почему колонию? Не колонию, а половинку! Ведь дочерние клетки останутся на месте своего рождения. Так возникнут двухцветные колонии. Но такое возможно лишь в том единственном случае, если нуклеоид несет на себе всю наследственную информацию и у мутантного гена нет "прикрытия" в виде неповрежденного аллеля, то есть бактерия гаплоидна.
Если бы хромосома бактерии имела близнеца, то комбинированные колонии появились бы только в случае, когда мутация повредила бы сразу два аллельных гена, что мало вероятно. Эти рассуждения оказались верны и для колоний, окрашенных на три четверти.
Опыты позволили Виткин заключить, что нуклеоид несет в себе всю наследственную информацию бактерии и что набор генов в нем один.
Доказательство гаплоидности бактерий было чрезвычайно важным для молекулярной генетики. Оно явилось той базой, на которой стало строиться ее здание, ибо обосновало важнейшее преимущество микробов как генетических объектов. Гаплоидны - это значит, что они не имеют двойного набора генов и каждый ген бактерии - в единственном числе. Представьте себе, что одного из генов на хромосоме бактерии поразила мутация. У него нет "прикрытия" в виде второго доминантного гена, и следствия мутации будут налицо уже через одно-два поколения. Это обстоятельство совместно с чрезвычайной скоростью размножения бактерий и возможностью изучать у них простые и единичные признаки позволило неслыханно ускорить процесс картирования хромосом бактерий.
Красоты, открывающиеся в царстве бактерий, превосходили все мечты охотников за генами. Уже тогда Ледерберг понял - в стране микробных гибридов картографы хромосом обоснуются надолго. И он не ошибся.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'