§ VII.1. Система Qβ-репликазы
История Qβ началась с утверждения, под которым - в то время, когда его высказал С. Спигелман [115], - подписались бы очень немногие из его коллег-биохимиков. Это утверждение состояло в том, что фаг Qβ использует специфичный фермент репликации, опознающий только РНК фага Qβ. В ответ на весь проявленный скептицизм Спигелман представил высокоочищенный и хорошо охарактеризованный фермент, способный катализировать воспроизведение инфекционной вирусной РНК в бесклеточной среде. Тот факт, что в бесклеточной системе действительно синтезировалась РНК, содержащая все инструкции, был продемонстрирован в следующем классическом эксперименте [116]. Раствор, не содержащий клеток, подвергали серийному разведению (оставляя на каждом этапе достаточно времени для воспроизведения), причем раствор в конечном разведении содержал менее 10-15 доли исходного числа природных фаговых матриц, т. е. практически не содержал ни одной такой матрицы, и все-таки этот образец обладал такой же инфекционностью, как и исходный. Более того, использование чувствительного к температуре мутанта исключало возможность того, чтобы источником информации служило что-либо, кроме молекул РНК. Носителем этой информации является только плюс-цепь; поэтому для воспроизведения инфекционных молекул необходим индукционный период, в течение которого должны накопиться комплементарные (неинфекционные) минус-цепи. Воспроизведение всей популяции может инициировать даже одна матричная молекула, и в таком случае оно приводит к образованию клона идентичных потомков. Это было показано в опыте (см. Р. Левисон и С. Спигелман [117]), в котором образец разбавляли и разливали в пробирки. Благодаря синхронной инициации синтеза можно было идентифицировать пробирки, получившие 0, 1, 2 или более молекул матрицы - в строгом согласии с распределением Пуассона. Опыты по "обрубанию" показали, что для узнавания ферментом необходимы участки, распределенные по всей длине молекулы, включая обе концевые группы: ни одну из двух половинок цепи по отдельности репликаза не узнавала. Так как плюс- и минус-цепи реплицируются при помощи одного и того же фермента, можно ожидать определенной симметрии в распределении комплементарных областей по длине цепи. Любые "внутренние" комплементарные участки одной цепи должны зеркально отражаться в комплементарной цепи, следовательно, они, вероятно, располагаются в 3'- и 5'-половинках симметрично.
Эту очень интересную структурную проблему можно решить путем анализа последовательности, который проводится (и частично завершен) в лаборатории Вейсмана [118]. Определением последовательности занималась также группа Спигелмана [119], которая показала, что 3'- и 5'-концы действительно в какой-то мере комплементарны. 5'-конец плюс-цепи имеет строение
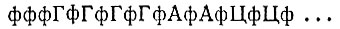
Минус-цепь тоже заканчивается группой фффГ на 5'-конце и имеет более длинную последовательность пуринов. Из этого следует, что 3'-конец плюс-цепи имеет область, комплементарную 5'-концу; из-за комплементарности с минус-цепью она должна содержать много урацила и цитозина и также кончаться цитозином.
Открытие системы Qβ может иметь как теоретическое, так и практическое значение. "Неожиданное структурное разнообразие и тонкость" (цитируется по С. Спигелману) отдельных молекул РНК делает понятным, каким образом силы отбора могли вмешаться во взаимодействие между нуклеиновыми кислотами и белками и направлять доклеточную эволюцию. В практических целях можно использовать специфичные участки узнавания: а) в сочетании с дегенерировавшей неинфекционной РНК, что может препятствовать фаговой инфекции; б) в качестве специфичных индукторов синтеза РНК в других, соответственным образом модифицированных системах.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'