§ VII.3. Количественные исследования отбора
Важные данные можно получить из количественных экспериментов по эволюции. Хотя в этом направлении еще предстоит много работы, некоторую информацию можно извлечь из уже опубликованных данных.
"Селективное преимущество" всегда относится к воспроизведению всего вида или ансамбля. Если скорость воспроизведения зависит от длины цепи, то просто утрата ненужной информации, приводящая к укорочению цепи - без увеличения "характеристической" скорости воспроизведения (которая относится к отдельной единице), - уже может оказаться "селективным преимуществом". Однако в условиях насыщения, когда скорость также становится независимой от концентрации матрицы, такая зависимость от длины цепи исчезнет. Термин "насыщение" означает, что практически все молекулы фермента Е связаны с матрицами Ii, т. е. 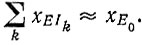 Такие условия явно обладают рядом преимуществ при кинетических исследованиях, и Спигелман использовал их в своих опытах. Если скорость воспроизведения не зависит от концентрации матриц, число матричных молекул (определяемое по включению 32Р) растет линейно во времени (рис. 26). Наличие периода индукции, заметного на рис. 26, указывает на различие в параметрах скорости и (или) связывания плюс- и минус-цепей. Если обозначить плюс- и минус-цепи
Такие условия явно обладают рядом преимуществ при кинетических исследованиях, и Спигелман использовал их в своих опытах. Если скорость воспроизведения не зависит от концентрации матриц, число матричных молекул (определяемое по включению 32Р) растет линейно во времени (рис. 26). Наличие периода индукции, заметного на рис. 26, указывает на различие в параметрах скорости и (или) связывания плюс- и минус-цепей. Если обозначить плюс- и минус-цепи 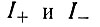 соответственно, а их концентрации
соответственно, а их концентрации 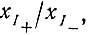 то кинетические уравнения для условий, указанных в подписи к рис. 26, будут иметь следующий вид:
то кинетические уравнения для условий, указанных в подписи к рис. 26, будут иметь следующий вид:
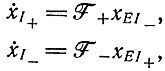 (VII.1)
(VII.1)где EI+ и EI- - комплексы фермент - матрица. Процедура, аналогичная выводу уравнения Михаэлиса-Ментен, приводит к выражению
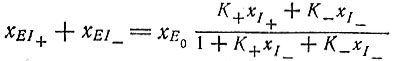 (VII.2)
(VII.2)(E0 - суммарное количество свободного и связанного фермента).
К+ и К- - константы стабильности комплексов фермент - матрица или их аналоги для стационарного состояния. Если затравкой служили одни только плюс-цепи, то начальный наклон относится только к включению в минус-цепи:
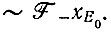 (VII.3)
(VII.3)
После накопления достаточно большого числа минус-цепей достигается постоянное "равновесное" отношение 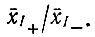
Принимая, что
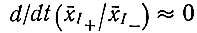
или
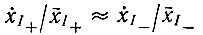 (VII.4)
(VII.4)и используя уравнение (VII.1), получаем
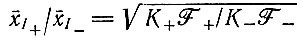 (VII.5)
(VII.5)или для конечного наклона, для тех условий, когда 32P включается и в плюс- и в минус-цепи:
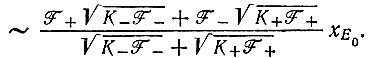 (VII.6)
(VII.6)
Последнее выражение при 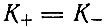 сводится к
сводится к
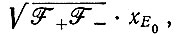
а для 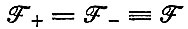 к
к
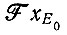
(ср. с результатами § IV.2, которые, однако, не относятся к ферментативному воспроизведению в области насыщения).
Из экспериментальных данных следует вывод, что плюс-цепь образуется быстрее, чем минус-цепь (каждая использует в качестве матрицы комплементарную цепь). Различие в наклонах можно объяснить различиями в значениях F и К (а не только К). Если различие обусловлено только параметром скорости, то F+ может быть в 100 раз больше, чем F-. Для количественного расчета необходимы, однако, более подробные экспериментальные данные. Конечный "вариант" (после 74 переносов) имеет более высокую среднюю скорость образования. Конечный наклон возрастает в 2,6 раза (по сравнению с конечным наклоном исходной РНК фага Qβ). Хотя имеются веские основания считать, что это изменение в основном обусловлено ростом параметров скорости, экспериментальные данные не позволяют точно вычислить отдельные F и К. Уменьшение периода индукции может быть обусловлено как ростом скоростей, так и присутствием плюс- и минус-цепи в начальной фазе. (Отметим, что периода индукции не должно быть, если обе цепи с самого начала представлены в своем "уравновешенном" соотношении.) Имеющиеся в настоящее время данные нельзя считать достаточными для обоснования вывода, что увеличение наклона в 2,6 раза (это определенно означает увеличение средней скорости копирования отдельных единиц) означает 15-кратное увеличение скорости воспроизведения индивидуальных "вариантных" молекул РНК (по сравнению с исходной РНК фага Qβ). Необходимо не только получить более детальные кинетические данные, но и доказать, что укорочение молекулы линейно связано с уменьшением времени ее полного воспроизведения, что маловероятно, если матрицу могут считывать одновременно несколько молекул фермента, и что должно зависеть также от концентраций. Тем не менее скорость воспроизведения отдельных молекул должна заметно расти, иначе включение 32Р - как показано на рис. 26 - не возрастало бы с числом переносов, несмотря на уменьшение времени инкубации в 4 раза.
Следует также подчеркнуть, что сделанные выше оценки относятся к самой простой модели взаимодействия фермент - матрица, предполагающей стационарное состояние, и были даны только для того, чтобы продемонстрировать возможности получения более полной информации - путем количественного исследования кинетики и механизмов.
Необходимо сделать еще одно замечание о механизме отбора. Основываясь на обсуждении в гл. II, можно было бы утверждать, что в области насыщения механизм отбора не действует. Процесс перестает быть "автокаталитическим", если x⋅i уже не пропорциональны xi, а являются постоянными. Однако это было бы верно только в том случае, если бы мы рассматривали репликацию идентичных матриц. Если же имеется несколько конкурирующих последовательностей Ii, каждая из которых может образовать комплекс с ферментом, характеризующийся константой стабильности
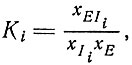 (VII.7)
(VII.7)то мы получим
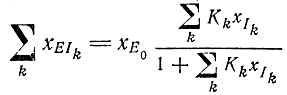 (VII.8)
(VII.8)или
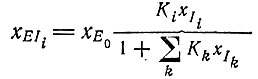
и
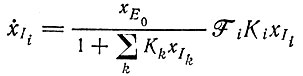 (VII.9)
(VII.9)
для 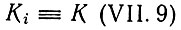 сводится к
сводится к
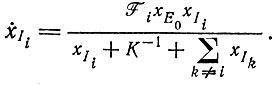 (VII.10)
(VII.10)
Физическая интерпретация следующая: даже в области насыщения, которая теперь определяется условием 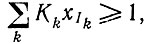 определенный мутант Ii, появляющийся в небольшом числе копий, воспроизводится в соответствии с механизмом экспоненциального отбора
определенный мутант Ii, появляющийся в небольшом числе копий, воспроизводится в соответствии с механизмом экспоненциального отбора 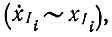 до тех пор пока концентрация отобранного вида не достигнет уровня, означающего доминирование:
до тех пор пока концентрация отобранного вида не достигнет уровня, означающего доминирование: 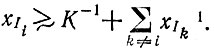 *
*
* (Для простоты здесь имеется в виду механизм "самоузнавания". Аналогичный основной результат получается и для комплементарного узнавания.)
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'