1.2.1* Открытие ядерной дРНК в эукариотических клетках
Роль клеточного ядра в синтезе РНК. В конце 1950-х годов в литературе шли жаркие споры о том, где происходит синтез РНК - в ядре или цитоплазме. Постепенно складывалось мнение, что синтез РНК практически полностью происходит в ядре. Это хорошо укладывалось в схему ДНК→РНК→белок, поскольку ядерная локализация, по крайней мере основной части ДНК, не вызывала сомнений. В пользу того, что синтез РНК идет в ядре, возможно на матрице ДНК, прежде всего говорили результаты опытов по включению радиоактивных предшественников в РНК ядра и цитоплазмы, которые были выполнены во многих лабораториях мира. Клетки инкубировали в течение короткого времени с радиоактивным предшественником, например уридином-[3Н], и затем смотрели на распределение включившейся метки между ядром и цитоплазмой с помощью так называемой авторадиографии. Срезы клеток покрывали фотоэмульсией и после выдержки и проявления видели, что при короткой инкубации клеток с меченым предшественником практически вся меченая РНК находится в ядре и лишь позднее начинает поступать в цитоплазму.
В то время в цитоплазме клеток выявляли два основных типа РНК- Это были высокомолекулярная, в основном рибосомная РНК (рРНК), состоящая из двух компонентов, и низкомолекулярная, в основном транспортная РНК (тРНК). Они хорошо разделялись при ультрацентрифугировании. Их скорости седиментации, описываемые так называемыми коэффициентами седиментации (S20, w). составляли 28S и 18S для рРНК и 4S для низкомолекулярной РНК. Это соответствовало длине ~5000, ~2000 и 70-100 нуклеотидов соответственно.
Нулеотидный состав цитоплазматической РНК определялся доминирующим компонентом, т. е. рРНК. Он отличался высоким содержанием гуаниловой (G) и цитидиловой (С) кислот. Отношение G + C/A (адениловая кислота)+ U (уридиловая кислота) составляло~1,6. В то же самое время в ДНК клеток животных преобладали А и Т; коэффициент специфичности G + C/A + T составлял около 0,7.
Было неясно, каким образом на матрице ДНК синтезируется РНК совсем другого нуклеотидного состава, т. е. возникало то же самое противоречие, что и при сравнении нуклеотидного состава ДНК и РНК бактерий, только еще более ярко выраженное.
Чтобы решить этот вопрос, предпринимались попытки изучить природу ядерной РНК, но это затруднялось отсутствием в то время методов выделения ядер, при которых РНК не деградировала бы.
Выделение недеградированной ядерной РНК1* [11 - 13]. В 1960 г. мы с В. Л. Мантьевой искали пути для выделения ядерной РНК в более или менее нативном состоянии, чтобы изучить ее свойства и выяснить природу. В это время широкое применение приобрел метод выделения нуклеиновых кислот с помощью обработки фенолом. После обработки клеток смесью фенола и воды (или солевого раствора) и разделения фаз центрифугированием РНК переходила в водный слой, а большая часть белков растворялась в феноле (рис. 1). В определенных условиях, если добавлять в водный раствор детергенты, то в него переходит и ДНК. Фенол не только сразу удалял значительную часть белков, но и ингибировал нуклеазы клетки, и это помогало получать неповрежденные нуклеиновые кислоты. Мы использовали этот метод для выделения РНК и ДНК.
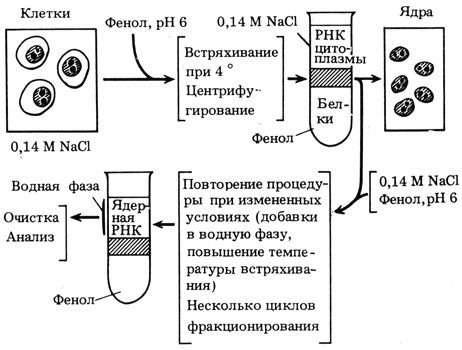
Рис. 1. Принципиальная схема метода фенольного фракционирования ядерной РНК и выделения в чистом виде дРНК (гяРНК). Схемы фракционирования, использованные при открытии дРНК (1961 г.): 0,14 М NaCl/фенол, рН 6,0° (3 раза); 5 % n-аминосалицилат/фенол, рН 6, 0° - дРНК; 0,14 М NaCl/фенол, рН 6, 60° - рРНК; при создании метода термического фенольного фракционирования (1962 г.): 0,14 М NaCl/фенол, рН 6, 0° (3 раза); 40° - рРНК; 55° - рРНК + дРНК; 65° - дРНК; при создании конечного варианта этого метода (1967 г.): 0,14 М NaCl/фенол, рН 6, 0° (3 раза); 40° - рРНК; 55° - рРНК+дРНК; 0,14 М NaCl+додецилсульфат/фенол, рН 6, 63° - дРНК; 85° - дРНК (по результатам, полученным В. Л. Мантьевой, В. Я. Арионом и соавт.)
После встряхивания клеток, суспендированных в 0,14 М NaCl с фенолом (рН 6), на холоду и последующего центрифугирования на границе между водной (верхней) и фенольной фазами образовывался слой нерастворимого материала, который содержал всю ДНК и сравнительно небольшую часть РНК и белков клетки. При этом основная часть клеточной РНК присутствовала в водной фазе, а белков - в фенольной. Из промежуточного слоя были взяты мазки, окрашены, и при их микроскопии неожиданно оказалось, что нерастворимый материал, или "интерфаза", содержит сохранившие свою форму клеточные ядра. В них были хорошо различимы даже ядрышки (см. рис. 1). Мы предположили, что "фенольные ядра" окажутся хорошим объектом для выделения из них ядерной РНК, так как клетки сразу обрабатывались фенолом, ингибирующим нуклеазы. После ряда неудачных попыток извлечь РНК из "фенольных ядер" я использовал обработку в той же системе 0,14 М NaCl - фенол (рН 6), но при повышенной температуре (65°). При этом в водную фазу перешла значительная часть РНК "фенольных ядер", которая затем была очищена путем ряда переосаждений, после чего начались опыты по ее характеристике.
Во-первых, было проведено исследование размеров полученной РНК с помощью аналитического ультрацентрифугирования в лаборатории А. С. Спирина (Институт биохимии им. А. Н. Баха АН СССР), который был заинтересован в изучении нашей РНК, поскольку все изучавшиеся им препараты высокомолекулярных клеточных РНК эукариот всегда содержали только два компонента рРНК и никаких других видимых фракций РНК. В наших препаратах ядерной РНК он надеялся увидеть что-либо необычное. Однако при ультрацентрифугировании опять выявились все те же два компонента рРНК с константами седиментации 28S и 18S. Правда, кроме них, был виден гетерогенный материал, делавший пики рРНК не очень красивыми. Однако наиболее простым объяснением его появления было предположение о частичной деградации и агрегации РНК в ходе ее выделения.
Открытие ядерной дРНК1* [13, 14]. Чтобы "закрыть тему" и написать статью о свойствах ядерной РНК, оставалось определить нуклеотидный состав РНК из "фенольных ядер" и доказать ее рибосомную природу. Я приготовил эту РНК в больших количествах, и В. Л. Мантьева начала определение ее нуклеотидного состава. Однако он оказался сильно отличным от состава рРНК. Отношение G+C/A+U составляло 1,1-1,2, а не 1,6, как в рибосомной. Мы подумали, что это результат методической ошибки и старались подогнать результат под предполагаемый, но первоначально полученный результат упорно воспроизводился (табл. 1).
Возникло противоречие между данными по ультрацентрифугированию (2 пика рРНК) и по нуклеотидному составу (резкое его отличие от рРНК). Тогда мы вспомнили о гетерогенном материале, выявлявшемся при ультрацентрифугировании наряду с 28S и 18S пиками, и подумали, что противоречие можно объяснить, если допустить существование двух компонентов в составе ядерной РНК, а именно рРНК и другой, еще неизвестной РНК с резко отличным нуклеотидным составом и гетерогенными размерами. Размытость пиков при ультрацентрифугировании могла зависеть от присутствия этой новой РНК, а не от деградации рРНК, как мы думали вначале.
Мы начали пробовать разные эмпирические способы фракционирования РНК "фенольных ядер". Несколько опытов не дали успеха. Наконец, в одном из опытов фенольные ядра вначале обработали смесью 5 %-ного n-аминосалицилата фенола, а затем, после центрифугирования, материал интерфазы 0,14 М NaCl - фенолом при 65°. При этом около половины РНК перешло вместе с ДНК в водную фазу во время первой экстракции, а другая часть - во время второй (см. табл. 1). Нуклеотидный состав второй фракции РНК оказался очень похож на рРНК (G + C/A-|--U~l,5), а у первой он резко отличался. Отношение G+C/A + U составляло ~0,8 т. е. было близко к таковому для ДНК (0,7).
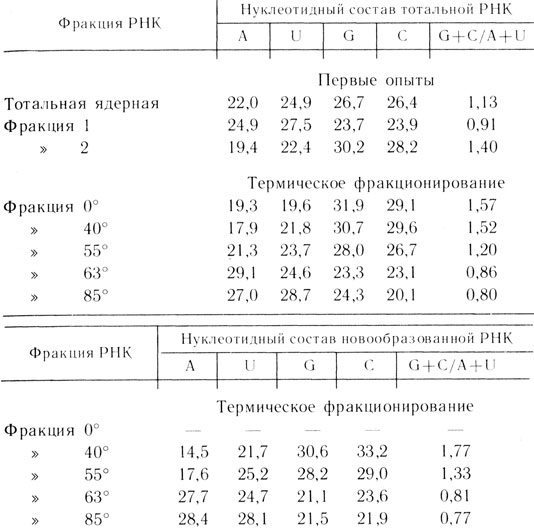
Таблица 1. Нуклеотидный состав фракций РНК асцитного рака Эрлиха,
Сразу стало ясно, что мы открыли новый тип РНК, а именно РНК, по нуклеотидному составу похожею на ДНК клетки. Мы назвали ее РНК AU-типа, а позднее использовали термин дРНК (ДНК-подобная РНК). Повторив этот опыт несколько раз, мы убедились в правильности заключения. Таким образом, как это часто бывает, открытие явилось следствием противоречия между ожидаемым тривиальным результатом и истинным ответом, полученным в эксперименте. Важно не пропускать такие чреватые открытиями ситуации, а искать другие объяснения и ставить опыты, позволяющие их проверить.
Шла весна 1961 г. Мы стали думать о природе РНК AU-типа, а также о публикации полученных данных. В это время стали появляться первые слухи о выявлении мРНК в клетках бактерий. Естественно было предположить, что и мы обнаружили мРНК, или информационную РНК животных клеток. Странным было лишь ее очень высокое содержание-около 1/3 всей РНК "фенольных ядер".
Первые сообщения о существовании РНК AU-типа мы опубликовали в работах [13, 14]. Данные об РНК AU-типа были доложены летом 1961 г. на V Международном биохимическом конгрессе в Москве (секционное сообщение), но на фоне сообщений об открытии генетического кода они прошли незамеченными. Ими заинтересовался лишь А. Мирский (США). Хотя в его собственном докладе ничего похожего не было, но весной 1962 г. он и его сотрудники сообщили о сходных с нашими результатах. Был использован другой метод фракционирования, который в отличие от нашего давал полностью деградированную РНК (4-6S). Дальнейшего развития его работа не имела.
Метод фенольного термического фракционирования 1* [16-18]. Как бы то ни было, начиная с осени 1961 г. мы с В. Л. Мантьевой начали систематическую работу по изучению ядерной AU-РНК, или дРНК. Первый метод нам не понравился прежде всего в силу своей недостаточно хорошей воспроизводимости. Первое, что пришло в голову,- это использовать для выделения РНК из "фенольных ядер" обработку при разных температурах. Постепенно повышая температуру (с интервалами в 5°), я получал разные фракции РНК, а В. Л. Мантьева анализировала их состав. При 40 и 45° в раствор выходила чистая рРНК (G+C/A+U = 1,5-1,6), при последующей обработке при 50 и 55°- РНК со смешанным нуклеотидным составом, а при 60 и 65° - чистая дРНК (G + C/A+U = 0,7-0,8) (см. табл. 1,2). Метод, по крайней мере на использованных типах клеток - асцитной карциноме Эрлиха мышей и печени крысы,- был простым и очень хорошо воспроизводимым. Работа была опубликована в 1962 г.
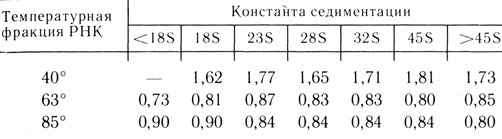
Таблица 2. Нуклеотидный состав фракций новообразованной РНК (G+C/A+U), полученных путем фенольного фракционирования и последующего гель-электрофореза (по результатам, полученным В. Я. Арионом и Г. Марковым)
Метод фенольного термического фракционирования (далее использовались лишь три ступени: 40, 55 и 65°, а позднее еще и 85°) давал возможность не только продемонстрировать существование ядерных дРНК и рРНК, но и нарабатывать их в миллиграммовых количествах, а следовательно, сделал возможным изучение их свойств.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'