6.1. Сплайсинг
Сплайсинг про-мРНК в клеточном ядре [242-247]. Центральным событием процессинга, естественно, является сплайсинг про-мРНК. К сегодняшнему дню усилиями многих групп (В. Келлер, ФРГ, Ф. Шарп, Т. Маниатис, Дж. Стейц, США, и др.) расшифрованы основные этапы сплайсинга. Удалось выделить ряд промежуточных компонентов и ряд необходимых для сплайсинга факторов, используя как клеточные, так и бесклеточные системы сплайсинга.
Первым этапом сплайсинга про-мРНК является разрыв фосфодиэфирной связи у 5'-конца интрона. Далее, освободившийся фосфорилированный 3'-конец интрона (pGpUpN...) переносится к участку, расположенному на расстоянии ~ 30 нуклеотидов от 3'-конца интрона, и присоединяется к 2'-гидроксильной группе аденозина. В этом месте в интроне располагается сильно вырожденная последовательность PyPyPuAPy... (Py - пиримидин, Pu - пуриннуклеотид). Образуется структура Типа "лассо". Третий этап - атака свободным 3'-гидроксилом экзона фосфодиэфирной связи у 3'-конца интрона. В результате образуется нормальная фосфодиэфирная связь между экзонами и вырезание интрона в виде лассо (рис. 41).
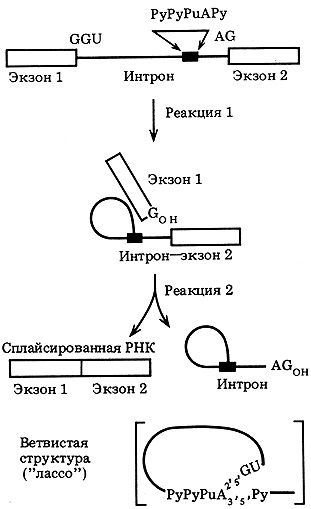
Рис. 41. Основные этапы реакции сплайсинга в гяРНК эукариот (схема) (по результатам, полученным Дж. Абельсоном и соавт.)
Сплайсинг, таким образом, - достаточно сложный процесс. Он осуществляется макромолекулярным комплексом, носящим название сплайсосомы. В состав последней входят рибонуклеосома и ряд белков. Кроме них, большая роль в сплайсинге принадлежит малым ядерным РНК (мяРНК), входящим в состав мяРНП.
Уже давно обнаружено, что последовательность у 5'-конца одной из мяРНК, называемой U1, комплементарна усредненной 9-нуклеотидной последовательности, расположенной на 5'-границе интрона. Действительно, мяРНК U1 необходима для сплайсинга. Инъекция в ядро антител к белкам РНП частиц, содержащих РНК U1, блокирует сплайсинг. Разрушение 5'-конца этой РНК также его подавляет, тогда как 3'-конец РНК Ш можно разрушить, сохранив активность последней. Более детальные исследования показали, что РНК U1 нужна для первого этапа сплайсинга (отщепления 5'-конца интрона) и, видимо, только для него. Для второго этапа, образования лассо, необходим другой тип мяРНК, который содержит мяРНК U2. Последняя, вероятно, может образовывать дуплекс с вовлекаемым в образование лассо участком интрона вблизи от А. Получены данные, что третий этап реакции осуществляется при участии еще одной мяРНК, а именно U5. Наконец, каким-то образом в процесс сплайсинга вовлечены еще мяРНК U4 и U6.
Итак, большая роль в реакции сплайсинга принадлежит мяРНК, причем последние участвуют в этом процессе через образование дуплексов со специфическими отрезками про-мРНК. Следовательно, мяРНК играют активную роль в узнавании мест сплайсинга. Из этого вытекают интересные возможности. Весьма вероятно, что существует еще серия мяРНК или вариантов U1 и U2, которые несколько отличаются от них по нуклеотидным последовательностям в местах узнавания. Тогда такие варианты мяРНК будут предпочтительно связываться с участками, отличными от канонических, и обеспечивать сплайсинг по другим местам. Таким образом, можно было бы объяснить тканеспецифический сплайсинг наличием разного набора мяРНК в разных тканях. Вопрос этот требует дальнейшего изучения.
Аутокаталитический сплайсинг [248-250]. Одной из главных сенсаций при изучении сплайсинга явилось открытие аутокаталитического сплайсинга у про-рРНК тетрахимены, а затем и в ряде других систем Т. Чех и соавт. (США) показали, что вырезание 400-нуклеотидного нитрона из про-рРНК тетрахимены идет при инкубации чистой РНК в присутствии ионов Mg и GMP. Последний можно заменить на GTP, GDP или просто гуанозин. Таким образом, была открыта возможность биокатализа при участии самой РНК, что имеет огромное биологическое значение.
Аутосплайсинг идет через следующие этапы (рис. 42): 1) 3'-гидроксил гуанозина атакует фосфодиэфирную связь (-UpA-) у 5'-конца интрона и присоединяется к нему через нормальную 3'-5' фосфодиэфирную связь (GpA-); 2) освободившийся 3х - ОН-конец (-U-ОН) экзона атакует фосфодиэфирную связь (-GpU-) у 3'-конца интрона, в результате чего экзоны соединяются (-UpU-); 3) освободившийся 3' - ОН-конец (-G-ОН) интрона атакует фосфодиэфирную связь (-UpA-) внутри интрона, что ведет к образованию из интрона двух цепей РНК: кольцевой (содержит -GpA-) и линейной (pGpA-...-U-ОН).
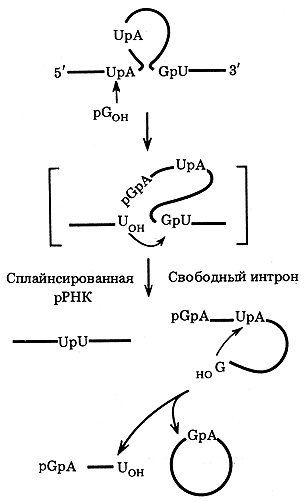
Рис. 42. Основные этапы самосплайсинга (автокаталитического сплайсинга) рРНК тетрахимены (схема) (по результатам, полученным Т. Чехом и соавт.)
Таким образом, сплайсинг двух экзонов и вырезание интрона в форме двух цепей РНК является результатом трех последовательных реакций переноса фосфодиэфирной связи. Очевидно, в РНК образуется такая вторичная структура, что, во-первых, определенные участки РНК сближаются, а во-вторых, снижается энергетический барьер для реакции переноса фосфодиэфирной связи. На самом деле, in vivo в реакции сплайсинга про-рРНК участвует также и белок, но его роль не каталитическая, а сводится к поддержанию оптимальной для сплайсинга конфигурации про-рРНК. Аутосплайсинг выявлен и в ряде других случаев. В митохондриях сплайсинг ряда классов РНК осуществляется путем РНК-катализа, а белкам опять-таки принадлежит лишь вспомогательная функция. Не исключено, что и в описанном выше сплайсинге про-мРНК роль мяРНК не является пассивной, но этот вопрос требует дальнейшего изучения.
Открытие РНК катализа на примере самосплайсинга легло в основу новых эволюционных теорий, согласно которым первичной формой живой материи была самовоспроизводимая молекула РНК- Сейчас описано еше несколько примеров биокатализа, осуществляемого РНК. Так, созревание некоторых тРНК у бактерий осуществляется РНКазой Р, в составе которой выявлена особая РНК. Оказалось, что именно эта РНК является ферментом, и она может сама, без белка, расщеплять молекулу РНК. Наконец, некоторые олигонуклеотиды могут катализировать полимеризацию нуклеозидтрифосфатов по своей матрице. Вероятно, с течением времени появятся еще примеры реакций, катализируемых не белком, а РНК. Здесь же я хотел бы лишь еще раз подчеркнуть особую роль РНК в организации сплайсинга, основного этапа процессинга про-мРНК.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'