2.3.1. Структура хроматина
2.3.1.1. Уникальная и повторяющаяся ДНК
Избыточность ДНК в геноме человека. Вскоре после того, как генетический код был расшифрован (в начале 60-х гг.), ученые пришли к выводу об избыточности ДНК в эукариотических клетках. По данным разных авторов, содержание ДНК в диплоидной клетке человека составляет примерно 7,3·10-12 г (размах от 6,6 до 8,0). Зная мол. массу оснований, можно подсчитать, что нуклеотидная пара А-Т (аденин - тимин) имеет массу 1,025·10-21 г, а нуклеотидная пара G-C (гуанин - цитозин) - 1,027·10-21 г. Следовательно, весь диплоидный набор содержит приблизительно 7,1·109 нуклеотидных пар:
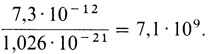
Если вся эта ДНК входит в состав структурных генов, кодирующих белки, а средний белок, подобно гемоглобину, состоит примерно из 150 аминокислот, то человеческий геном должен содержать примерно 6-7 млн. генов [1338; 1339]. В настоящее время известно, что эта цифра завышена примерно на два порядка. "Информативная" (кодирующая) ДНК чередуется с последовательностями, которые не транслируются в аминокислотные последовательности. Некоторые из них имеют какие-то специфические функции, для других функции до сих пор не обнаружены.
Факты, свидетельствующие об избыточности ДНК в клетках эукариот, были известны и раньше. Например, при изучении гигантских хромосом Drosophila и Chironomus оказалось, что диски в этих хромосомах имеют среднюю длину 20000-50000 нуклеотидных пар (20-50 т.п.н.). С другой стороны, данные генетического анализа свидетельствуют о том, что один диск (+ междиск) в норме содержит только один ген [1042]. Прямой анализ генома человека, однако, нуждается в новых методах.
Повторяющаяся ДНК [1317; 509; 409]. Большую роль в развитии представлений о структуре генома сыграло открытие того, что ДНК высших организмов содержит большую фракцию повторяющихся последовательностей. Выделенную из клеток высокомолекулярную ДНК можно фрагментировать на отрезки примерно одинаковой длины, а затем такие короткие двухцепочечные структуры денатурировать, т. е. разделить на одноцепочечные с помощью нагревания в солевом растворе. В таком растворе одноцепочечные фрагменты могут свободно перемещаться и случайно сталкиваться один с другим. При резком понижении температуры они, встречаясь с комплементарными партнерами, будут формировать двойные спирали ДНК. В этом состоит простой метод установления комплементарности цепей ДНК.
Если бактериальную ДНК подвергнуть тепловой денатурации указанным образом, а затем идентифицировать фракцию реассоциировавшей после отжига двухцепочечной ДНК по исходной концентрации молекул C0 и времени реакции (t), то получается линейная зависимость (на логарифмическом графике этому соответствуют S-образная кривая, C0t-кривая) (рис. 2.78). Если провести такой эксперимент с фрагментами человеческой ДНК длиной примерно в 600 пар оснований, то кривая будет совершенно другая. Сразу же после начала отжига обнаруживается небольшой процент реассоциировавшей двухцепочечной ДНК. Крутой наклон кривой показывает, что следующая фракция ДНК отжигается примерно в 50000 раз быстрее, чем бактериальная ДНК; еще одна фракция ДНК отжигается быстрее бактериальной в 10-1000 раз. Остальная ДНК (≈50%) характеризуется такой же кинетикой, как и бактериальная. Эти данные можно объяснять следующим образом: небольшая часть ДНК человека имеет области, в которых комплементарные последовательности располагаются на одной и той же цепи, но в обратном порядке (палиндром). Эта ДНК может реассоциировать очень быстро, просто складываясь вместе. Другая фракция содержит повторяющиеся последовательности, которые реассоциируют, образуя двухцепочечную ДНК; в данном случае скорость реассоциации зависит от числа идентичных (или почти идентичных) повторов. Наконец, имеются еще уникальные последовательности ДНК (единичные копии), кинетика реассоциации которых сходна с таковой для бактериальной ДНК (рис. 2.78).
![Рис. 2.78. Кинетика отжига фрагментов ДНК разной длины у бактерии и человека. Указана доля (%) двухцепочечной, реассоциированной ДНК для разных концентраций продукта ДНК (C0) и времени (t). Пунктирная S-образная кривая соответствует бактериальной ДНК и характерна для уникальной фракции. Точечная кривая-профиль реассоциации фрагментов ДНК человека длиной в 600 оснований. Можно выделить четыре класса: 9% имеют неизмеримо быструю скорость отжига; 22% характеризуются C0t1/2=10-2; 12,5% - C0tl/2=1,0 и 51,2% - C0tl/2=495. C0tl/2=10-2 означает, что отжиг происходит примерно в 50000 раз быстрее, чем при C0tl/2=495. Нижняя кривая показывает кинетику реакции фрагментов длиной 1,3 т.п.н. Они реассоциируют намного быстрее. Это означает, что большинство сегментов содержит повторяющиеся последовательности. Только около 10% ДНК ведет себя как уникальная. (Данные Schmid, Deininger, 1975; рисунок из [499])](pic/000097.jpg)
Рис. 2.78. Кинетика отжига фрагментов ДНК разной длины у бактерии и человека. Указана доля (%) двухцепочечной, реассоциированной ДНК для разных концентраций продукта ДНК (C0) и времени (t). Пунктирная S-образная кривая соответствует бактериальной ДНК и характерна для уникальной фракции. Точечная кривая-профиль реассоциации фрагментов ДНК человека длиной в 600 оснований. Можно выделить четыре класса: 9% имеют неизмеримо быструю скорость отжига; 22% характеризуются C0t1/2=10-2; 12,5% - C0tl/2=1,0 и 51,2% - C0tl/2=495. C0tl/2=10-2 означает, что отжиг происходит примерно в 50000 раз быстрее, чем при C0tl/2=495. Нижняя кривая показывает кинетику реакции фрагментов длиной 1,3 т.п.н. Они реассоциируют намного быстрее. Это означает, что большинство сегментов содержит повторяющиеся последовательности. Только около 10% ДНК ведет себя как уникальная. (Данные Schmid, Deininger, 1975; рисунок из [499])
Как уникальные и повторяющиеся последовательности ДНК расположены относительно друг друга? В разных работах показано, что несколько больше 50% ДНК генома человека представлено уникальными фрагментами длиной около 2 т.п.н. Они распределены в основном между умеренно повторяющимися последовательностями, длина которых составляет 0,3 т.п.н. Многие из этих повторяющихся последовательностей весьма сходны друг с другом. Кроме того, высокоповторяющиеся последовательности ДНК, образованные миллионами копий коротких олигонуклеотидов, были обнаружены в таких специфических районах, как центромерная область или длинное плечо Y-хромосомы. Высокоповторяющаяся ДНК часто демонстрирует индивидуальные количественные и качественные различия, не влияющие, однако, на фенотип. Уникальная ДНК включает в себя структурные гены, но лишь небольшая часть этой ДНК представлена структурными генами. Описанная топология последовательностей очень широко распространена и наблюдается даже у весьма отдаленных видов, таких, как млекопитающие, амфибии, гастроподы и даже жгутиковые [509]. Широкое распространение относительно стабильного паттерна предполагает какую-то важную его функцию, которая, к сожалению, пока не выявлена. У некоторых видов, например у Drosophila melanogaster и Chironomus tentans, подобное распределение коротких последовательностей ДНК не обнаружено.
Повторяющиеся последовательности ДНК со специфическими функциями. Некоторые умеренно повторяющиеся последовательности содержат гены, необходимые всем клеткам в каждой фазе индивидуального развития (рибосомной РНК, гистонов, транспортной РНК). Как правило, гены рибосомной РНК (рРНК) являются частью района ядрышкового организатора, а само ядрышко содержит пул рРНК. У человека районы ядрышковых организаторов расположены в коротком плече акроцентрических хромосом (13-15; 21; 22). Для определения числа генов рРНК у человека был использован метод гибридизации РНК - ДНК in vitro [1021; 1022]. По соотношению объема фракции ДНК, гибридизующейся с рРНК, с общим содержанием ДНК в ядрах клеток человека, было определено среднее число копий рибосомных генов в диплоидной клетке. Оно оказалось равным приблизительно 416-443.
Мультигенное семейство, образованное многочисленными генами вариабельных участков иммуноглобулинов (разд. 4.4), включает столь большое количество копий, что соответствующие последовательности ДНК можно отнести к классу умеренно повторяющихся. В разд. 2.3.6.7 описываются другие мультигенные семейства, часть из которых может входить в повторяющуюся фракцию.
Сателлитная ДНК. Многие виды ДНК, особенно относящиеся к фракциям высокоповторяющихся последовательностей, характеризуются как сателлитная ДНК. При центрифугировании фрагментированной ДНК в градиенте плотности хлористого цезия выявляется основная полоса или пик. По обе стороны от основного пика часто видны маленькие пики. Соответствующая им ДНК и называется сателлитной. Количество и локализация пиков сателлитной ДНК видоспецифичны (рис. 2.79). Локализация пиков в градиенте плотности хлористого цезия определяется нуклеотидным составом последовательностей. Отдельный пик может стать заметным только в том случае, если состав этой фракции отличается от состава основной фракции ДНК. В хромосомах сателлитная ДНК обычно соответствует конститутивному гетерохроматину. У человека она находится также вне центромерной области в Y-xpoмосоме и в хромосомах 1, 9 и 16. Она состоит из коротких высокоповторяющихся последовательностей, которые могут быть представлены несколькими миллионами копий. (Сателлитную ДНК не следует путать с сателлитными районами акроцентрических хромосом. Использование одного и того же термина является неудачным совпадением.) Сравнение фракций сателлитной ДНК человека и других видов, особенно высших обезьян, весьма важно для понимания эволюции человека.
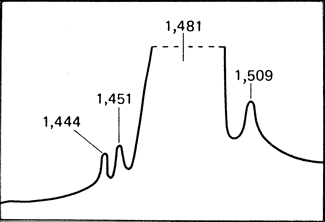
Рис. 2.79. Сателлитная ДНК человека: аналитическое ультрацентрифугирование тотальной плацентарной ДНК в градиенте плотности сульфата цезия в присутствии ионов серебра свидетельствует о наличии сателлитов I (1,444), II (1,451), и III (1,509). (По Miklos and John, Amer. J. Hum. Genet., 31, p. 266, 1979.)
Функция сателлитной ДНК неизвестна и является предметом дискуссий. Например, предполагают, что сателлитная ДНК участвует в распознавании гомологичных хромосом во время мейотической конъюгации или модулирует некоторые регуляторные функции генов. Пока нет убедительных доказательств в пользу той или иной гипотезы. Однако исследования на дрозофиле свидетельствуют о влиянии сателлитной ДНК на кроссинговер [437]. Открытие сателлитной ДНК удивило цитогенетиков тем, что она оказалась локализованной в той части хроматина, которая по данным микроскопического анализа уже многие десятилетия идентифицировалась как гетерохроматин. Относительно недавних открытых "мини-сателлитных" последовательностей ДНК см. разд. 2.3.2.7.
2.3.1.2. Гетерохроматин
Определение и свойства. Термин "гетерохроматин" был предложен Хейтцем (Heitz, 1928) [469]. Он писал: "У P. (Pellia)epiphylla (мох) некоторые участки пяти из девяти хромосом ведут себя по-разному. В телофазе в отличие от остальных участков этих пяти и других четырех хромосом они продолжают оставаться видимыми, даже могут наблюдаться в молодых интерфазных ядрах, а также в ядрах полностью сформировавшихся клеток". Сохранение конденсированного состояния в интерфазе является основной характеристикой гетерохроматина [313]. Позднее были обнаружены другие особенности. Например, репликация ДНК во время S-фазы происходит в гетерохроматиновых сегментах намного позже, чем в эухроматиновых. Обычно различают два класса гетерохроматина: конститутивный и факультативный. У человека факультативная фракция представлена инактивированной Х-хромосомой у женщин и мужчин, которые несут дополнительные Х-хромосомы (разд. 2.2.3.3).
Гетероморфизмы: функция и отношения с сателлитной ДНК [410]. Существует значительная межиндивидуальная изменчивость гетерохроматина (раздел. 2.1.2.3), превышающая изменчивость эухроматической части генома. Такие варианты называют "гетероморфизмами". Кроме тех районов, о которых говорилось ранее (вторичные перетяжки хромосом 1, 9, 16), обнаружен гетероморфизм в центромерных и спутничных районах акроцентрических хромосом. В медико-генетической практике этот гетероморфизм используется, например, для установления материнского или отцовского происхождения хромосом у больных с геномными мутациями, такими, как синдром Дауна (разд. 5.1.2.3), или в случае спорного отцовства. В течение многих лет было распространено представление о том, что в конститутивном гетерохроматине нет классических менделевских генов, но большинство исследователей не хочет признавать отсутствие у конститутивного хроматина какой-либо функции. Напротив, предполагают, что у него много функций. Возможно, например, что он влияет на стабилизацию структуры хроматина и выполняет роль "телохранителя", защищая генетически значимые последовательности ДНК эухроматических районов от внешних воздействий [385].
Высказанные соображения наталкивают на мысль, что явления, описанные в классической цитогенетике и положенные в основу понятия о гетерохроматине, сродни тем фактам, которые были открыты совсем недавно и легли в основу таких понятий, как высокоповторяющаяся и сателлитная ДНК (хотя эти недавние результаты получены с помощью совершенно иных экспериментальных подходов). И сателлитная, и высокоповторяющаяся ДНК, и гетерохроматин расположены главным образом вблизи центромеры, но могут обнаруживаться и в других районах некоторых хромосом (1, 9, 16, Y). В них отсутствуют известные структурные гены, но они не полностью идентичны: например, небольшие количества сателлитной ДНК были обнаружены при помощи гибридизации in situ (разд. 2.3.2.3 и 7.2.2) вне центромерного конститутивного гетерохроматина. Функциональный смысл такой локализации остается неясным.
2.3.1.3. Нуклеосомная структура хроматина [1172; 421]
Химический состав хроматина. Кроме ДНК хромосома содержит много разных белков. Вместе с двухцепочечной ДНК эти белки образуют хроматин. Среди них больше всего гистонов - положительно заряженных щелочных белков с молекулярной массой около 10000-20000. Их можно разделить на пять классов (H1, Н2А, Н2В, Н3 и Н4). Другие, так называемые негистоновые, белки представлены в очень небольших варьирующих количествах. Негистоновая фракция гетерогенна, в нее входит, например, много ферментов.
Нуклеосомы [1172]. Хроматиновая нить состоит из повторяющихся единиц - нуклеосом, представляющих собой набор гистоновых молекул, ассоциированных примерно с 200 парами нуклеотидов ДНК. Набор гистонов в таких единицах состоит из любых двух белков четырех типов: Н2А, Н2В, Н3 и Н4. Они свернуты как глобулы, образуя цилиндр. ДНК-вый компонент нуклеосомы имеет две части: "сердцевину" (или кор - от англ. core) из 140 пар нуклеотидов (п. н.) и "связующее звено" (или линкер - от англ. linker). Длина линкера варьирует от 15 до 100 п. н. в зависимости от типа клетки. Такие линкеры, очевидно, связывают нуклеосомы друг с другом. Гистон H1, который почти вдвое длиннее, чем другие гистоны, отвечает за целостность нуклеосомной структуры. При удалении H1 (что легко сделать в эксперименте) цепочка становится менее плотно упакованной (рис. 2.80, 2.81). На одну нуклеосому приходится только одна молекула гистона H1. ДНК закручена вокруг набора из девяти гистонов; вместе они образуют сферическую частицу с диаметром около 100 Å. Такие частицы лежат, плотно прилегая друг к другу, вдоль хроматинового волокна. Точно не известно, каким образом ДНК связана с гистонами, однако ясно, что структура двойной спирали при этом явно не нарушена. Исследования, проведенные с помощью метода гибридизации ДНК - РНК (см. разд. 2.3.2.3), указывают на то, что в нуклеосомах встречается широкий спектр функционально различных последовательностей ДНК, от уникальных до повторяющихся, активно транскрибирующихся и таких, которые встречаются в конститутивном гетерохроматине. Вероятно, вся хромосомная ДНК эукариотической клетки упакована в нуклеосомы. Доказательства нуклеосомной структуры опираются на три типа данных: электронно-микроскопическое исследование хроматина обнаруживает цепочки частиц, анализ дифракции рентгеновских лучей показывает наличие в хроматине повторяющихся единиц, и наконец, ферментативный гидролиз хроматина микрококковыми нуклеазами позволяет изолировать отдельные нуклеосомы. Возникновение понятия нуклеосомы стимулировало проведение новых экспериментов в различных направлениях, результаты которых в конце концов подтвердили существование нуклеосом и помогли установить свойства этих структур.
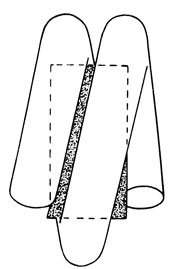
Рис. 2.80. Схематическое изображение отдельной нуклеосомы
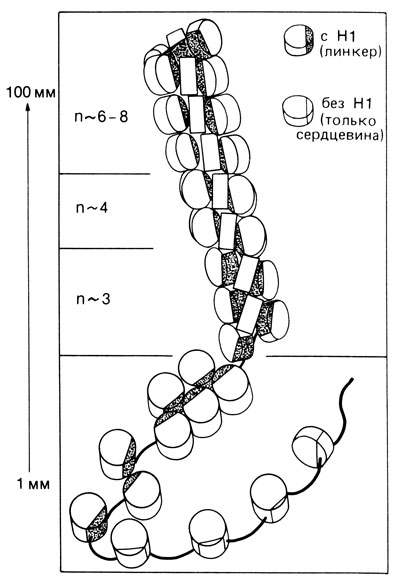
Рис. 2.81. Схематическое изображение нуклеосомной структуры хроматина: в опытах in vivo структура зависит от концентрации соли. При 100 ммоль NaCl на одном витке хроматиновой нити умещается 6-8 нуклеосом (вверху). При снижении концентрации соли остается только 3-4 нуклеосомы на виток (в центре). В отсутствие соли нуклеосомы мало контактируют одна с другой. (По Küppers, Molekulare Genetik, 3rd, ed., 1982.)
2.3.1.4. Интеграция хроматиновых волокон в хромосомную структуру
Интерфаза. Интерфазная хромосома представляет собой элементарную фибриллу, состоящую из нуклеосом, соединенных линкерами. Эта фибрилла пронизывает не все ядро, а лишь определенные его области. Возможно, однако, что транскрибируемые хромосомные сегменты распространяются до центра ядра. В норме хроматин сильно спирализован. Относительно точного числа уровней спирализации еще ведется дискуссия. Волокна, соответствующие возрастающим порядкам спирализации, можно описать следующим образом [1042]:

Митотические и мейотические хромосомы. Как видно из этой таблицы, хромосомы в митозе и в мейозе обнаруживают значительно большую степень спирализации, чем в интерфазе (разд. 2.1.2). Рисунок их сегментации обсуждался в разд. 2.1.2.3. Число субсегментов, которые можно идентифицировать в составе сегментов, зависит от степени конденсации хромосомы (от митотической профазы до метафазы) и качества окрашивания. Это особенно отчетливо можно продемонстрировать при помощи метода преждевременной конденсации хромосом. Верхний предел задается числом хромомер 30000-100000 нуклеотидных пар в длину (см. ниже [201а]). Учитывая, что число нуклеотидных пар на гаплоидный геном приблизительно равно 3,5·109, а число сегментов, видимых даже в лучших препаратах, не превышает ≈2000 (разд. 2.1.2.3), можно сделать вывод, что нет даже близкого приближения к такому уровню разрешения. Хромосомные сегменты выявляются и во время ранних фаз мейоза.
Изучение рисунка репликации митотических хромосом показало, что ДНК темных G-сегментов (идентичных светлым R-сегментам и, как правило, ярко флуореоцирующим Q-сегментам) реплицируется обычно во второй половине S-фазы. Отдельный сегмент в прометафазной хромосоме является, по-видимому, единицей репликации (которая сама состоит из многих репликонов). Видимо, репликация начинается в одно и то же время. Предполагалось, что такая организация единицы репликации может иметь какое-то функциональное значение. Эти единицы содержат много высокоповторяющихся и нетранскрибируемых последовательностей ДНК. Количество видимых сегментов зависит от степени конденсации хромосом, как показано на рис. 2.82. В полностью деспирализованной хромосоме каждая функциональная единица, содержащая повторяющиеся, некодирующие участки и уникальные транскрибирующиеся участки, в идеальном случае могла бы распознаваться как структура, состоящая из G-сегмента вместе с R-сегментом. Из этого следует, что R-сегменты должны иметь большую плотность генов, чем G-сегменты и яркие Q-сегменты. В геноме человека такие районы с выраженными R-сегментами найдены в участках 3p, 6p, 11q, 12q, 17q и 19 (p или q). При изучении сцепления (разд. 3.4.3) действительно именно в этих районах оказалось больше генов, чем должно быть при случайном распределении. Кроме того, количество диагностируемых абортов, обусловленных трисомией по этим районам, меньше ожидаемого, следовательно, такая аномалия кариотипа приводит к очень ранней и потому невыявляемой гибели плода [1481]. Существует представление, согласно которому хромосома в метафазе состоит из чередующихся "областей сжатия" (вероятно, идентичных темным G-сегментам) и тех областей, в которых при определенных условиях могут образоваться петли [535]. Районы хромосомы, называемые в классической цитогенетике эухроматическими и гетерохроматическими, вероятно, по тонкой структуре друг от друга отличаются.
![Рис.2.82. Формирование паттерна хромосомной сегментации (G-сегменты) путем скручивания хроматидных нитей, которые состоят из слабоокрашенных (обогащенных эухроматином), а также из темных (гетерохроматиновых) областей. Обратите внимание, что число видимых G-сегментов уменьшается с увеличением плотности спирализации [201а]](pic/000101.jpg)
Рис.2.82. Формирование паттерна хромосомной сегментации (G-сегменты) путем скручивания хроматидных нитей, которые состоят из слабоокрашенных (обогащенных эухроматином), а также из темных (гетерохроматиновых) областей. Обратите внимание, что число видимых G-сегментов уменьшается с увеличением плотности спирализации [201а]
2.3.1.5. Интегральная модель структуры хромосомы
Эти данные вместе с результатами молекулярно-биологических исследований (см. ниже) позволяют сформулировать интегральную модель хромосомы: она состоит из единственной двойной спирали ДНК, объединенной с гистонами в нуклеосомы. Некоторые районы этой двойной спирали представлены в основном повторяющимися последовательностями, высокоповторяющиеся копии сателлитной ДНК могут быть рассеяны по геному. Участки, богатые повторяющимися последовательностями (в первую очередь в центромерной области и во вторичных перетяжках), обнаруживают признаки конститутивного гетерохроматина. Заметим, однако, что преобладающими в молекуле ДНК являются все-таки уникальные последовательности длиной в 2000 (и больше) нуклеотидных пар. Они рассеяны между мало и умеренно повторяющимися. При исследовании уникальных сегментов методами классической цитогенетики оказывается, что они обнаруживают свойства эухроматина и именно в них при определенных условиях выявляются петли большей или меньшей протяженности. Транскрибирующиеся последовательности ДНК (собственно "гены", см. разд. 2.3.5) локализованы преимущественно в этих уникальных районах, которые соответствуют светлым G-сегментам и темным R-сегментам. Особые последовательности, кодирующие рРНК, локализованы в районах ядрышкового организатора. Описанные структуры можно изучить детально в подходящих для этого клетках (например, в больших ооцитах амфибий). В этих же клетках можно наблюдать транскрипцию [440, 535].
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'