3.5.4. Группы крови: Rh-комплекс, неравновесие по сцеплению
История. В 1939 г. Левин и Стетсон [762] исследовали сыворотку крови женщины, которая родила мертвый плод и в анамнезе которой имело место переливание крови мужа, совместимой по ABO группе. При этом ими были обнаружены особые антитела. Позже Левин и Стетсон показали, что из 1010 образцов крови только 21 дал отрицательную реакцию с этими антителами. Выявленные антитела никакой связи с системами групп крови ABO, MN и P не имели.
В 1940 г. Ландштейнер и Винер [753] при иммунизации кроликов эритроцитами макака-резуса получили сыворотку, которая агглютинировала эритроциты 39 из 45 особей. При сравнении этих антител с антителами, обнаруженными Левином и Стетсоном, авторы пришли к выводу, что в обоих случаях реакция происходит с одним и тем же антигеном. В дальнейшем оказалось, что это не совсем так. В настоящее время антиген, открытый с помощью истинного анти-резус-антитела, называется LW - в честь Ландштейнера и Винера, а Rh-типирование у человека всегда проводится с сывороткой человеческого происхождения, как это было сделано в работе Левина и Стетсона. Последующее изложение вопроса относится только к реакциям с этими антителами человека.
Огромная практическая важность системы Rh стала очевидной, когда была установлена связь между этими антителами и несчастными случаями при переливании крови. Кроме того, стало понятным, что именно резус-несовместимость матери и плода является причиной эритробластоза плода и гемолитической болезни новорожденных. Оказалось, что эритроциты примерно 85% всех представителей белой расы дают положительные реакции с анти-Rh-сыворотками. Семейными исследованиями было установлено, что Rh-положительные индивиды являются гомозиготами Rh/Rh или гетерозиготами Rh/rh, тогда как rh-отрицательные индивиды - это гомозиготы rh/rh.
В 1941 г. Винер открыл другие антитела, которые реагировали с эритроцитами 70% всех индивидов и отличались от основного фактора Rh (Rh′ по Винеру). Третий родственный фактор был открыт в 1943 г. В семейно-популяционных исследованиях выявлены все возможные комбинации этих трех факторов, причем наследовались совместно именно комбинации. Винер выдвинул гипотезу, согласно которой эти серологические "факторы" являются "агглютиногенами" и что каждый из них детерминируется одним аллелем из серии множественных аллелей одного гена. Эта описательная гипотеза настолько неконкретна, что на ее основе можно было объяснить все, в том числе и позже открытые факты. Для того чтобы судить о внутренней структуре Rh-локуса, по мнению большинства исследователей, необходимо было провести биохимический анализ.
Гипотеза Фишера о тесно сцепленных локусах. В 1943 г. Фишер сформулировал более конкретную гипотезу. В то время удалось выявить еще одно антитело, анти-Hr, и Фишер, анализируя подготовленные Рейсом полные таблицы серологических данных, обнаружил, что Rh′- и Hr-факторы комплементарны. У каждого человека в крови присутствуют либо антиген Rh′, либо Hr, либо оба антигена. Индивид, имеющий оба антигена, никогда не передает их вместе одному потомку, т. е. ребенок всегда получает только один антиген из двух. Для объяснения этих фактов Фишер предложил модель, согласно которой пара аллелей определяет один из двух антигенов. Эта пара была названа C/c. Аналогично была постулирована дополнительная пара аллелей D/d для исходных антигенов Rh+ и rh-, а также третья пара аллелей для уже открытого тогда третьего серологического фактора. Кроме того, чтобы согласовать генетические данные о наследовании всех трех факторов, постулировалось наличие тесного сцепления между этими тремя локусами.
Гипотеза Фишера предполагала открытие двух недостающих (комплементарных D и E) антигенов d и e. Это предсказание подтвердилось для антигена e, но не для d. По-видимому, этот хромосомный район не содержит то "нечто", что приводит к образованию антител. В развитии данной гипотезы Фишер сделал важный шаг вперед. В британской популяции наиболее частыми были три класса комплексов Rh-генов (рис. 3.32, 3.33). По мнению Фишера, редкие комбинации появляются вследствие изредка происходящего кроссинговера.
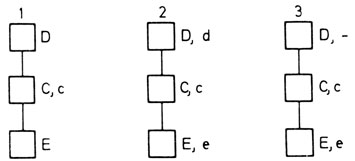
Рис. 3.32. Гипотетическая структура Rh-комплекса. 1. На основе данных, известных к 1941 г. 2. Антигены, предсказанные Фишером и Рейсом. 3. Уже открытые антигены (антиген d пока не обнаружен)
![Рис. 3.33. Предполагаемое образование трех редких Rh-гаплотипов из более частых вследствие кроссинговера (Race, Sanger [166]). Каждая диаграмма предполагает самостоятельное событие кроссинговера](pic/000199.jpg)
Рис. 3.33. Предполагаемое образование трех редких Rh-гаплотипов из более частых вследствие кроссинговера (Race, Sanger [166]). Каждая диаграмма предполагает самостоятельное событие кроссинговера
Действительно, все четыре комбинации, относящиеся к редким классам, могут возникнуть в результате кроссинговера между более частыми комбинациями, но для CdE это не так. Для появления этой комбинации необходим двойной кроссинговер. Следовательно, гипотеза объясняет, почему комбинация CdE так редка. Возможно и другое объяснение. При каждом кроссинговере, приводящем к возникновению Cde, CDE или cdE, должна возникать также и комбинация cDe. Отсюда следует, что суммарная частота первых трех комбинаций должна быть равна частоте cDe. Действительно, найденные частоты были такими: cDe - 0,0257 и Cde + cdE + CDE - 0,0241 (среди негров, однако, частота cDe была выше).
Кроме того, Фишер предположил, что три указанных локуса расположены в последовательности D-C-E, поскольку комбинация cdE, которая возникает вследствие кроссинговера между локусами D и E в генотипе cDE/cde, встречается намного чаще относительно этого генотипа, чем комбинация CDE относительно генотипа CDe/cDE (кроссинговер между C и E).
Подтверждение и предварительная интерпретация порядка расположения генов. За 30 лет, прошедших с тех пор, как Фишер выдвинул свою гипотезу, было сделано много новых наблюдений. Наиболее важным для решения вопроса о порядке расположения генов было выявление комбинированных антигенов, например се. Существование этого составного антигена, по-видимому, не противоречит последовательности D-C-E, тогда как составные антигены, предполагающие тесное сцепление между D/d и E/e, не были обнаружены. Гипотеза Фишера поставила два вопроса. 1. Если вследствие кроссинговера формируются иногда редкие комбинации из более частых, то в семейных исследованиях должны обнаруживаться случаи кроссинговера. Действительно, имелось сообщение об одной такой семье [896]: у отца с генотипом CDe/cde и матери с генотипом cde/cde было четверо детей cde/cde и трое - CDe/cde, что находится в полном соответствии с генетической теорией. Однако шестой (в порядке рождения) ребенок имел генотип Cde/cde. Этот факт можно было бы объяснить тем, что ребенок внебрачный. Однако такое объяснение кажется малоправдоподобным, если исходить из данных по другим группам крови и сывороточным факторам, а также учитывая принадлежность этой семьи к секте с особо строгими нравами. Однако других семей, подобных этой, обнаружено не было. Вполне вероятно, что многие исследователи просто не станут учитывать такой атипичный случай, поскольку заподозрят здесь методическую ошибку.
2. Какова должна быть структура Rh-локуса(ов) в свете достижений молекулярной генетики? Имеются две принципиальные возможности:
а) Rh-комплекс - это один цистрон с многими мутационными сайтами. Мутационные изменения выражаются в антигенных различиях;
б) Rh-комплекс состоит из нескольких тесно сцепленных цистронов (возможно, трех), и основные антигены отражают генетическую изменчивость по этим цистронам. В отсутствие каких-либо неопровержимых биохимических данных этот вопрос остается неразрешенным. Определенные выводы можно сделать на основании цис-транс-теста. Поскольку составной антиген се обнаруживается только в цис-положении CE/ce, но не в транс-положении Ce/cE, Рейс и Сэнгер (1969) [846] высказали гипотезу, согласно которой C/c и E/e относятся к одному функциональному гену.
За последние десятилетия накоплено множество фактов, которые свидетельствуют о том, что не только в структурном, но и в количественном отношении экспрессия Rh-фактора находится под строгим генетическим контролем. Розенфельд и соавт. (1973) [860] попытались обобщить все имеющиеся данные на основе новой модели структуры Rh-локуса. Согласно этой модели, Rh-локус состоит из нескольких областей (структурных генов), несущих информацию о мембранных полипептидах. Эти области находятся под контролем общего оператора или промотора, который регулирует количественную экспрессию, возможно, благодаря нескольким операторным районам, приближенным к единственному структурному гену. Эта модель объединяет фишеровскую концепцию с более поздними результатами молекулярной биологии. Однако биохимические доказательства модели пока отсутствуют.
Неравновесие по сцеплению. В процессе обоснования гипотезы о наследовании Rh-комплекса Фишер разработал еще одну концепцию: неравновесие по сцеплению. Обычно сцепление не приводит к ассоциации признаков в популяции (разд. 3.4.1). Даже если в начальной популяции фазы сцепления распределены не случайно, то многократно повторяющийся кроссинговер будет рандомизировать комбинации аллелей в группе сцепления, и в конце концов фазы притяжения и отталкивания для двух сцепленных локусов будут встречаться в популяции с одинаковой частотой. Это случай равновесия по сцеплению. Однако если в начальной популяции существует отклонение от равновесия, то время, за которое оно будет достигнуто, зависит от степени сцепления: чем теснее сцепление, тем больше требуется времени для достижения равновесия. И оно никогда не будет достигнуто, если определенные комбинации аллелей определяют сниженную приспособленность.
Правда, селективный недостаток некоторых аллельных комбинаций Rh-комплекса, способных обусловить снижение их частоты, до сих пор еще не продемонстрирован: отбор работает против гетерозигот (разд. 6.2), но это не означает, что общее снижение приспособленности никогда не существовало или никогда впредь не будет иметь убедительного объяснения в терминах истории популяции. Отвечая на некоторые вопросы, гипотеза Фишера в свою очередь поставила ряд других. Сама по себе концепция неравновесия по сцеплению остается важной в генетическом анализе полиморфизма ДНК (разд. 6.1) и главного комплекса гистосовместимости.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'