1.2.2. Нарушенное расщепление диплоидов
Описанный выше анализ сцепленного наследования генов основывается на определенном цитологическом механизме - механизме мейоза у диплоидов, при котором хромосомы двух наборов закономерно попарно конъюгируют, обмениваются участками, расходятся.
У некоторых штаммов грибов этот обычно четко работающий механизм может нарушаться - в одном ядре может быть не две гомологичные хромосомы, а больше, хромосомы могут иметь перестройки, изменяющие характер их конъюгации, обмен участками может быть подавлен. Наконец, часть продуктов мейоза может оказаться нежизнеспособной в результате проявления летальных генов или эффектов хромосомных перестроек. Некоторые из упомянутых нарушений препятствуют генетическому анализу, затрудняют его, другие - напротив, могут быть использованы в таком анализе.
Первое, что мы в связи с этим рассмотрим, - последствия летальности части аскоспор. Многие природные и особенно производственные штаммы дрожжей показывают низкую жизнеспособность аскоспор. Гибриды этих штаммов и лабораторных линий также не дают большого числа асков с четырьмя жизнеспособными спорами. Если в таких случаях удается все-таки собрать достаточно представительную выборку асков с двумя выжив-шими спорами, то рекомендовано проводить так называемый диадный анализ [20], который в отличие от анализа случайной выборки аскоспор позволяет определять сцепление не только генов, но и генов и центромеров.
Если идет расщепление по двум генам А/а и B/b и среди выживших спор по каждому из них наблюдается соотношение аллелей 1:1, то возможно появление диад 10 типов (табл. 1.9). Из табл. 1.9 можно вывести формулы для определения числа асков Р-, N- и Т-типа, которые дали наблюдаемые частоты диад разных типов Х1 - Х10 из общего их числа n:
Отсюда с помощью обычных формул можно рассчитать значение сцепления между генами и определить, сцеплены ли они с центромерами.
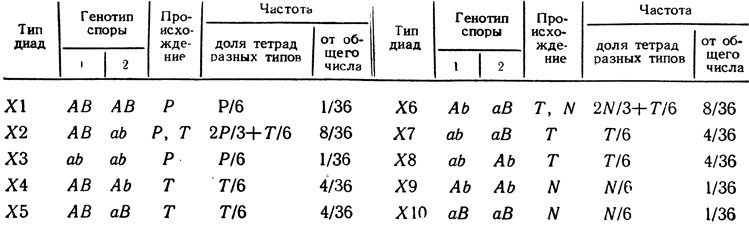
Таблица 1.9. Типы диад, их происхождение и частота
Рассмотрим теперь последствия подавления рекомбинации. Как указывалось, расхождению гомологичных хромосом в мейозе предшествует обмен участками между ними. Как следствие, гены, локализованные в разных хромосомах, показывают 50 % рекомбинации; расположенные в одной хромосоме (в зависимости от их расстояния) - от 0 до 50 %, т. е. достаточно удаленные ведут себя как несцепленные и принадлежность их к одной хромосоме не может быть обнаружена. Подавление мейотической рекомбинации могло бы облегчить анализ, так как при этом все гены одной группы сцепления передавались бы единым блоком и было бы легко разбить весь набор генов на отдельные группы сцепления.
Подавление рекомбинации может быть следствием мутаций. Одна такая мутация найдена у дрожжей-сахаромицетов: рецессивная мутация spoil (VIII хромосома), как оказалось, подавляет в гомозиготном состоянии мейотическую рекомбинацию. Для ее использования в генетическом анализе при определении локализации генов анализируемую мутацию соединяют с мутацией spoil и такой штамм скрещивают с тестерами, несущими, помимо spoil, маркеры всех групп сцепления. В споровом потомстве полученных гибридов маркеры разных хромосом рекомбинируют свободно, картирумая мутация не даст рекомбинантов лишь с маркером, лежащим в хромосоме, гомологичной той, в которой располагается эта мутация [24].
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'