20. Сцепленные гены
Гены, расположенные в одной и той же хромосомной паре, называются "сцепленными". Сцепленные гены не подчиняются второму закону Менделя; они не дают свободной рекомбинации друг с другом. У норки мутантные гены черных и кареглазых пастелей являются сцепленными; такими же, разумеется, являются их нормальные аллеломорфы. Сцепленные с полом гены сцеплены с определяющими пол генами в хромосоме; естественно, они также сцеплены и друг с другом.
У человека мутантные гены дальтонизма и гемофилии связаны с полом (см. гл. 15) и, следовательно, сцеплены друг с другом. Случается, что оба мутантных гена могут оказаться в одной семье. Такие семьи бывают двух различных типов.
Тип I. В этих семьях два мутантных гена соединились в итоге брака между людьми, несущими тот или иной ген. Гены, таким образом, пребывают в различных хромосомах; выражаясь языком генетики, они сцеплены в процессе "отталкивания" (рис. 62, I, первый ряд).
Тип II. В этих семьях мутация, приводящая к одной из таких ненормальностей, например гемофилии, совершается в хромосоме, которая уже несет другой мутантный ген. Оба мутантных гена находятся в одной и той же X-хромосоме; они сцеплены в процессе "притяжения" (см. рис. 62, II, первый ряд).
Большинство женщин в обоих типах семей будут фенотипически нормальны, но многие окажутся гетерозиготными по одному или обоим мутантным генам (см. рис. 62, второй ряд), и эти женщины могут иметь больных сыновей. Так как сын наследует только одну X-хромосому матери, больные сыновья гетерозигот I типа страдают или гемофилией, или дальтонизмом, но не имеют оба заболевания одновременно (см. рис. 62, третий ряд), в то время как больные сыновья гетерозигот II типа имеют оба заболевания одновременно. Однако могут быть и исключения. Они представлены в четвертом ряду рис. 62.
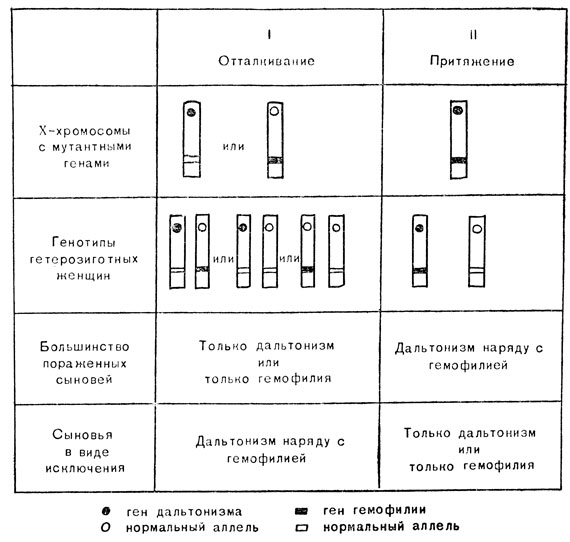
Рис. 62. Связь между сцепленными с полом генами цветной слепоты и гемофилии
Изредка женщина, несущая ген дальтонизма в одной X-хромосоме и ген гемофилии в другой (тип I), может родить сына, страдающего обоими заболеваниями; или же женщина, несущая гены обоих заболеваний в одной X-хромосоме (тип II), может родить сына, страдающего только одним из этих заболеваний.
Нетрудно заметить, что эти исключения аналогичны таковым у детенышей паломино-норки, соединивших в одной хромосоме два гена, которые у их родителей находились в различных хромосомах-партнерах. Во всех этих случаях мутантный ген, по-видимому, перемещается из одной хромосомы к ее партнеру. Этот процесс называется кроссинговером; он допускает ограниченное число рекомбинаций между генами, которые не подчиняются второму закону Менделя.
Явление кроссинговера породило большое количество экспериментов и предположений на протяжении десяти лет, но до сих пор оно остается непонятым до конца. Однако последствия кроссинговера хорошо известны и в любом случае могут быть предсказаны в такой же степени, как и последствия законов Менделя.
На рис. 63 показаны результаты кроссинговера для двух хромосом, вернее, для генов, находящихся в них.
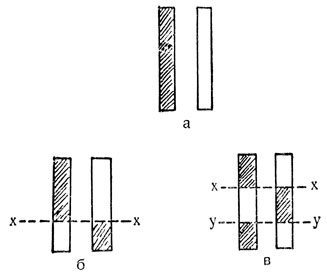
Рис. 63. Кроссинговер: а - пара хромосом; б - кроссинговер в одной точке: обмен концевыми кусками; в - кроссинговер в двух точках: обмен средними кусками
В исходной паре хромосом (a) один из партнеров заштрихован для отличия его от другого, б и в представляют собой два варианта многочисленных типов кроссинговера, которые могут наблюдаться у пары а. В обоих случаях произошел обмен кусками между хромосомами-партнерами. В случае б обмен получился концевыми кусками, и для этого потребовалась только одна точка (x) обмена; в случае в произошел обмен средними кусками, и для этого потребовались две точки (x и y).
Наиболее важная черта кроссинговера - точное соответствие между точками обмена у хромосом-партнеров. Если бы не было столь точного соответствия, то хромосомы-партнеры вскоре перестали бы быть равными по длине; более того, число генов в них перестало бы быть одинаковым, и хромосомы с очень большим или малым количеством генов могли попасть в круговорот, а это привело бы к появлению уродств и смерти.
Если, например (рис. 64), точка обмена в одной из хромосом попадет между 4-м и 5-м генами, а в другой - между 6-м и 7-м генами, то в результате обмена концевыми кусками получится одна хромосома с утерей 5-го и 6-го генов, а хромосома-партнер будет иметь эти гены в двойном количестве.
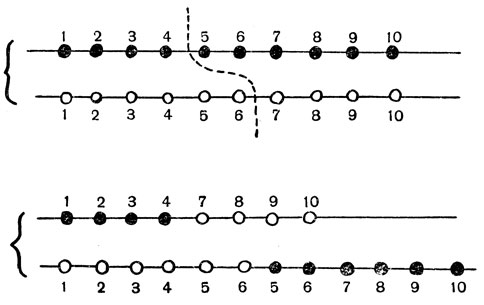
Рис. 64. Необходимость точности при кроссинговере. Если точки обмена у двух хромосом неточно подогнаны, то в получающихся хромосомах или будут отсутствовать гены (верхняя хромосома в нижнем ряду), или хромосомы будут нести двойное число генов (нижняя хромосома в нижнем ряду)
Наиболее распространена точка зрения, что кроссинговер происходит в начале мейоза, когда хромосомы-партнеры не только тесно и точно соприкасаются, но и обвиваются одна вокруг другой; вследствие напряжения от скручивания может произойти разрыв хромосом в идентичных точках и соединение вновь с кусками между партнерами. Высказываются и другие предположения, но до настоящего времени нет единой точки зрения на этот вопрос.
Обмен кусками между хромосомами-партнерами легко объясняет кроссинговер в его генетическом проявлении независимо от того, какой механизм лежит в его основе.
На рис. 65 показано, как можно объяснить рождение сына с дальтонизмом и гемофилией у женщины, которая несет гены этих аномалий в противоположных хромосомах (I), ли сына только с одной аномалией у женщины, имеющей оба гена в одной и той же хромосоме (II).
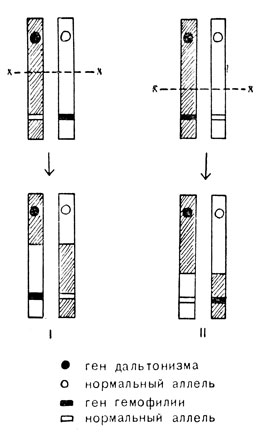
Рис. 65. Кроссинговер между генами дальтонизма и гемофилии
Если локализация точек обмена определяется случайностью, нужно ожидать, что кроссинговер между генами, значительно удаленными друг от друга в хромосоме, будет происходить чаще, чем между близко расположенными генами; это действительно так и происходит. Когда два гена расположены друг к другу очень близко, вероятность того, что точка обмена попадает между ними, невелика, и кроссинговер наблюдается редко. Чем больше расстояние между двумя генами, тем больше вероятность того, что точка обмена расположится между ними, и тем выше встречаемость кроссинговера. Два гена могут комбинироваться так же свободно, как если бы они находились в различных парах хромосом, в тех случаях, когда расстояние между ними больше какой-то определенной величины.
Исторически и логически это положение ставит повозку впереди лошади. Именно в результате открытия генетического сцепления стало возможным привязать гены к хромосомным парам, тогда как открытие кроссинговера позволило измерить расстояние между генами в единицах частоты встречаемости кроссинговера. Сказать, что два гена расположены друг от друга на расстоянии 10 единиц кроссинговера, проще и короче, чем говорить, что гетерозигота этих двух генов образует 10% кроссинговерных гамет.
Когда пара хромосом несет несколько или много известных генов, эксперименты с кроссинговером можно использовать для получения хромосомной "карты", которая показывает расположение генов и их относительные расстояния друг от друга.
У Drosophila melanogaster гены красных глаз (St, рецессивный), торчащих щетинок (Sb, доминантный) и изогнутых крыльев (Cu, рецессивный) расположены в третьей хромосоме, одной из двух длинных аутосом. Расстояние по кроссинговеру между St и Sb составляет 14%, а между Cu и Sb равно 8%. Одних этих данных недостаточно для того, чтобы представить себе расположение трех генов, так как они могут находиться в двух различных положениях (рис. 66, I и II). Однако когда станет известно, что расстояние по кроссинговеру между St и Cu равно 6, а между Cu и Sb равно 8, то порядок расположения генов St, Cu и Sb устанавливается точно (см. рис. 66, III). Таким же методом на карте может быть нанесено положение и других генов. В настоящее время карта этой хромосомы дает расположение более 150 генов. Тот факт, что гены всегда можно нанести на карту таким приемом, является доказательством их линейного расположения вдоль хромосомы. Если бы расположение было другим, например некоторые гены выступали из хромосомы в боковые ветви, то расстояние между тремя генами нельзя было бы всегда выражать так, что одно расстояние представляет собой сумму двух других (рис. 67). В действительности оно обычно оказывается несколько меньшим этой суммы; но эта второстепенная деталь может быть объяснена, и на этом здесь не стоит останавливаться.
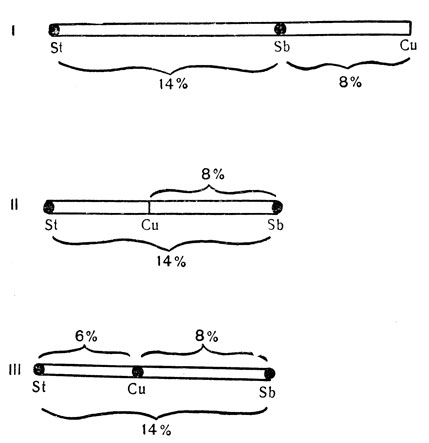
Рис. 66. Как гены 'наносятся' на карту. Расстояние между St и Sb равняется 14% кроссинговера; расстояние между St и Сu равно 8% кроссинговера; расстояние между St и Сu равно 6% кроссинговера. Следовательно, порядок расположения генов будет таким, как показано в III строке
Для селекционера-животновода значение кроссинговера заключается в возможности рекомбинаций, т. е. соединения или разделения сцепленных генов.
Можно ли этого достигнуть легко и экономично, зависит от расстояния между генами, которые интересуют селекционера. Когда гены достаточно далеко удалены друг от друга, сцепление не является препятствием для рекомбинаций. Когда сцепленные гены значительно сближены, чтобы можно было уловить сцепленность, селекционер сначала должен выяснить, достаточно ли у него возможностей для осуществления задуманного плана разведения.
Специалист, желающий скрестить породу мышей с розовыми глазами1 и породу шиншилла, чтобы получить породу мышей цвета шиншилла с розовыми глазами, может достигнуть этого без особого труда и затрат; так как расстояние кроссинговера между генами розовых глаз и мехом цвета шиншилла составляет около 15%, поэтому гетерозиготы, несущие эти гены, при отталкивании образуют около 15% гамет, несущих эти гены в протяжении. С другой стороны, любитель, разводящий мышей и желающий получить породу мышей с ослабленной окраской шерсти и нормальными ушами от породы со светлой окраской шерсти и короткими ушами, должен прежде обратиться к руководству по генетике мышей, чтобы выяснить, будет ли он в состоянии это осуществить, так как гены светлой шерсти и коротких ушей сцеплены очень тесно друг с другом, и если мышь несет оба гена в сцеплении в одной хромосоме и оба нормальных аллеломорфа в другой, то только 1 гамета приблизительно на 1000 будет нести ген светлой шерсти без гена коротких ушей.
1 (Ген розовых глаз отличается от гена альбиноса, обусловливающего белый цвет меха и розовый цвет глаз, и является фактически аллеломорфом гена шиншиллы)
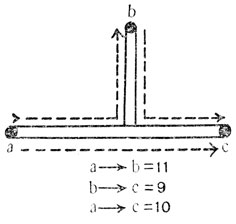
Рис. 67. Рисунок доказывает, что хромосома не имеет боковых ветвей. Если бы b лежал в конце боковой ветви, расстояние от a до c не являлось бы суммой расстояний от а до b и от b до c
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'