Глава 2. Как разгадали структуру генов
Итак, гены оказались состоящими из ДНК - вещества, которое биохимики, как им казалось, хорошо знали.
Строение ДНК. ДНК - это полимер. Полимерными молекулами химики называют длинные цепочки, состоящие из одинаковых (гомополимеры) или разных (гетеро- полимеры) звеньев - мономеров. В основе мономера ДНК - молекула углевода - пентоза. Обычно в молекулах углеводов отношение атомов кислорода и водорода такое же, как в воде, - 1:2, поэтому общая формула углеводов СnН2nОn.
Углевод ДНК - исключение из правила: в нем не хватает одного атома кислорода (С5Н10О4), поэтому его назвали обескислороженной рибозой - дезоксирибозой.
К каждому остатку дезоксирибозы в ДНК присоединено одно гетероциклическое (азотистое) основание. Их в норме четыре: аденин, гуанин, тимин, цитозин (рис. 2).
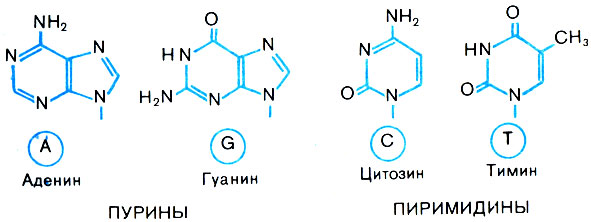
Рис. 2. Строение азотистых оснований ДНК
Гетероциклическими химики называют кольца, слагаемые, кроме углерода, и другими атомами, в данном случае азотом. Соединение основания с сахаром называют нуклеозидом.
Сокращенно основания и соответствующие мономеры ДНК обозначаются латинскими буквами A, G, С и Т. (В популярной литературе и учебниках их обозначают русскими буквами - соответственно А, Г, Ц, Т.) Углевод в ДНК также циклический: связь через атом кислорода образует пятиугольную молекулу. Атомы углерода в ней химики перенумеровали - 1, 2, 3, 4, 5, начиная с того, к которому присоединено основание.
Что сшивает отдельные нуклеозиды ДНК в единый полимер? Химики установили, что сшивки - остатки фосфорной кислоты Н3Р04. Такие связи называют фосфодиэфирными. Фосфодиэфирная связь в природных ДНК возникает между третьим и пятым атомами углерода в соседних пятичленных циклах пентозы, поэтому она называется 3′-5′-связью.
Нетрудно понять, что на одном конце образующегося полимера остается свободным 3′-атом углерода, а на другом - 5′. Эти концы так и называют 3′-и 5′-концами (рис. 3).
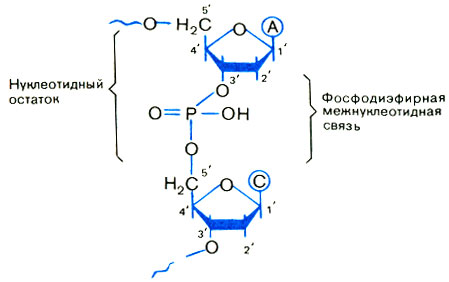
Рис. 3. Фрагмент цепи ДНК. Коротко он записывается - АС -
Добавим, что нуклеозид с остатком фосфорной кислоты называется нуклеотидом (а точнее, дезоксирибонуклеотидом). Это и есть отдельное звено нуклеотидной цепи ДНК.
Раньше полагали, что ДНК состоит из монотонно повторяющихся блоков - A, G, С, Т. Казалось, что полимер такого простого строения, такая "скучная" и однообразно устроенная молекула не может быть носителем разнообразных наследственных свойств. Но в конце 40-х годов австрийский биохимик Э. Чаргафф показал, что процент оснований G+C может варьировать в широких пределах, изменяясь от одного вида к другому. Значит, в природе может существовать бесконечное количество вариантов ДНК. Если длина полинуклеотида хотя бы тысяча звеньев, то количество вариантов 41000 (это число с шестью тысячами нулей!). Известно, что только двумя символами, например точкой и тире в азбуке Морзе или же нулем и единицей в программах ЭВМ, можно записать какой угодно большой объем информации. Так что кажущаяся простота строения ДНК не помеха для того, чтобы этот полимер выступил в роли "вещества наследственности".
Из анализов Э. Чаргаффа следовало, что в ДНК любого организма количество А всегда равняется количеству Т, а количество G - количеству С. Лишь потом из этого правила нашлись исключения - ДНК некоторых вирусов. Для подавляющего большинства организмов правила Чаргаффа строго соблюдались. Почему это так, стало ясно после работы Д. Уотсона и Ф. Крика, сделавших в 1953 г. одно из величайших открытий в истории естествознания. Они установили, что молекула ДНК - двойная спираль, в которой две полинуклеотидные оси закручены друг вокруг друга и вокруг общей оси (рис. 4).
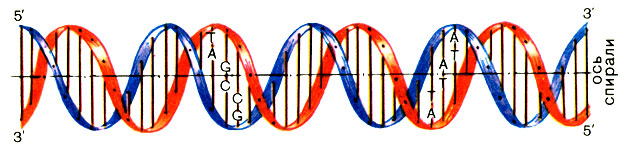
Рис. 4. Строение молекулы ДНК
Цепи ДНК антипараллельны: двигаясь мысленно вдоль длинной оси двойной спирали, в одной цепи мы будем проходить межнуклеотидные связи в направлении 3′-5′, а в другой - 5′-3′. Представьте двух змей, скрутившихся в спираль - голова одной к хвосту другой (рис. 5).

Рис. 5
Допустим, что голова - 5′, а хвост - 3′. Спираль ДНК обычно закручена вправо, как штопор или болт с правой резьбой. Но в некоторых случаях ДНК образует и левую спираль, названную Z-формой.
Что же связывает единичные цепи в двойной спирали? Д. Уотсон и Ф. Крик установили, что антипараллельные последовательности в ней образуют так называемые комплементарные пары. G в одной цепи соединяется с С в другой, так же как А с Т водородными связями, о которых вы имеете представление из школьного курса химии. Эти связи с участием электроположительного водорода (протона) и какого-либо электроотрицательного центра (кислорода, азота). Для существования жизни в земном варианте водородные связи чуть ли не важнее всех прочих: ведь вода кипит при 100° С именно потому, что молекулы ее связаны друг с другом в легкоразрушающиеся глобулы (ассоциаты) связями между кислородными и водородными ядрами. Исчезни эти связи - и все океаны на Земле вскипели бы.
Водородные связи гораздо слабее ковалентных; поэтому двойная спираль распадается на комплементарные половинки, если раствор ДНК подщелочить или нагреть до 100° С. Этот процесс называется денатурацией, он обратим. Если понизить рН раствора или понизить температуру, половинки ДНК, сталкиваясь в хаотическом тепловом движении, в конце концов находят друг друга, пары А-Т и G-С восстанавливаются, и снова возникают двойные спирали. Конечно, скорость этого процесса (ренатурации) зависит от концентрации раствора и от разнообразия последовательностей ДНК. Грубо говоря, чем больше разных генов "денатурируется", тем медленнее восстанавливаются двойные спирали ДНК. Образно говоря, чем разнообразнее груда обуви в прихожей, тем дольше приходится искать галошу под пару.
Так как в двойной спирали ДНК G всегда в паре с С, а А с Т, понятно, почему строго соблюдается правило Чаргаффа. У тех вирусов, которые не подчинялись этому правилу, ДНК оказалась одноцепочечной.
Возможность репликации заложена в самой ДНК. Нетрудно понять, что цепи в ДНК не только антипараллельны, но и комплементарны (А против Т и G против С). Зная последовательность нуклеотидов в одной цепи, мы легко восстановим последовательность в другой.
А теперь расплетем двойную спираль ДНК, разделим слагающие ее цепи и, пользуясь правилом комплементарности, построим на них новые (рис. 6). Две дочерние молекулы, которые мы получили, неотличимы от материнской. Вот и ответ на вопрос: как гены удваиваются в числе при делении клеток. Добавим, что водородные связи, возникающие между парами G и С, А и Т, самые устойчивые. Поэтому процесс удвоения, редупликации, генов очень помехоустойчив, ошибки возникают редко. Это и обеспечивает наследственность, преемственность поколений. Иначе дети бы не походили на родителей, в каждом поколении появлялась бы "ни мышонок, ни лягушка, а неведома зверушка".
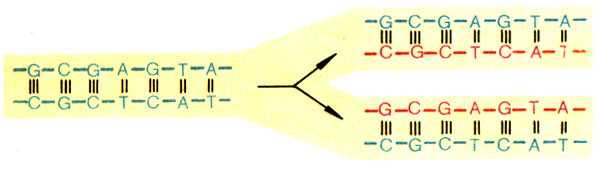
Рис. 6. Построение новых нитей ДНК на основе правила комплементарности
Редупликация ДНК идет с затратой энергии и ускоряется в клетках целым комплексом белковых молекул- катализаторов - ферментов, главный из которых называется ДНК-полимеразой. После расплетения двойной спирали ДНК этот фермент последовательно наращивает на полинуклеотидных цепях исходной молекулы ДНК (их называют также матричными цепями) комплементарные им цепи.
Обратите внимание: образование двух новых комплементарных цепей идет в противоположных направлениях. Это происходит потому, что матричные цепи антипараллельны, а ДНК-полимераза наращивает цепь только от 5′-конца к 3′-концу. То, что изображено на рисунке, называют обычно репликативной вилкой (рис. 7). Ее можно разглядеть в электронный микроскоп.
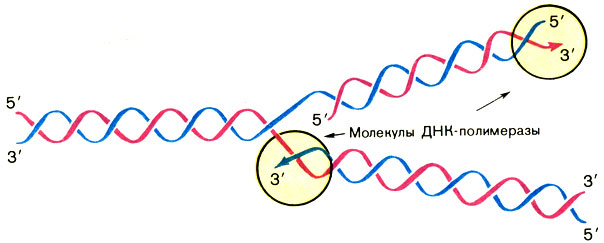
Рис. 7. Репликативная вилка
С ДНК-полимеразой, равно как и с принципом комплементарности оснований, мы еще не раз встретимся на страницах этой книги. А сейчас обратимся к вопросу о том, как реализуется содержащаяся в ДНК наследственная информация.
От ДНК к РНК и обратно. Известно, что основу жизни представляют белки. Функции их в клетках очень многообразны. Сокращается мышца, работает мозг, борется организм с попавшими в него бактериями или вирусами, переваривается в желудке и кишечнике пища, насыщается кровь кислородом, светится светляк, бактерия разъедает камень - во всех разнообразных процессах главную роль играют белки. Одного не "умеют" молекулы белков - размножаться. На них не распространяется принцип редупликации, свойственный генам. Но вся информация, необходимая для постройки белковых молекул, содержится в генах. Если в ядре нет соответствующего гена, организм не будет синтезировать определенный белок. Поэтому уже в 30-е годы было сформулировано правило: один ген - один белок.
У высших организмов - эукариот - белки образуются в цитоплазме клетки, а гены (ДНК) скрыты за оболочкой ядра. Поэтому ДНК непосредственно не может быть матрицей для биосинтеза белка. Эту роль выполняет другая нуклеиновая кислота - рибонуклеиновая (РНК). РНК отличается от ДНК по многим свойствам, но по составу различается незначительно. Вместо дезоксирибоз РНК слагают рибозы (это "нормальные" углеводы - С5Н10О5). РНК вместо тимина содержит урацил (U), отличающийся от тимина только тем, что у него нет метильной группы СН3. Однако РНК практически всегда состоит из одной полинуклеотидной цепи, хотя может образовывать комплементарную двойную спираль с породившей ее однонитчатой ДНК. Впрочем, внутри цепи РНК часто встречаются участки с комплементарными последовательностями нуклеотидов, отчего они скручиваются в двунитчатые "шпильки". Здесь также образуются комплементарные пары оснований, только А-Т заменена на А-U (рис. 8).
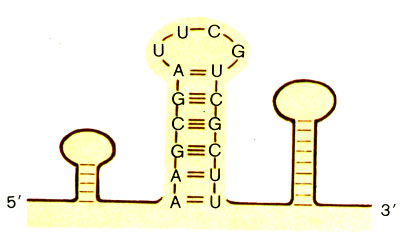
Рис. 8. 'Шпильки', образуемые полинуклеотидной цепью РНК
РНК синтезируется на ДНК, главную роль здесь играет фермент РНК-полимераза.
Матрицей для синтеза служит одна из цепей двуспиральной ДНК, которая в этом участке расплетается. Молекулярные биологи назвали этот процесс транскрипцией (переписыванием). Транскрипция ДНК строго следует принципу комплементарности, и поэтому образующаяся полинуклеотидная цепь РНК в точности комплементарна матричной нити ДНК и в принципе может образовать с ней идеальную двойную спираль. В следующих главах мы встретимся с такими ДНК - РНК-комплексами. Их называют ДНК - РНК-гибридами, а процесс их образования - ДНК - РНК-гибридизацией. В генной инженерии это свойство часто используют. Далеко не вся ДНК в ядре может образовывать транскрипты. Нетранскрибируемая ДНК служит для каких-то других целей, нам до конца еще неясных. Возможно, она организует структуру хромосом или выполняет обязанности регулятора.
Оказалось, что процесс транскрипции обратим. Известен фермент, синтезирующий на РНК как на матрице полинуклеотидную цепь ДНК. Эта вновь синтезированная ДНК - точная копия той матричной цепи гена, с которой была считана (транскрибирована) РНК. Фермент назвали РНК - зависимой ДНК-полимеразой или, проще, обратной транскриптазой или ревертазой (от слова реверс - обратный ход). Он обнаруживается в составе вирусов, вызывающих перерождение нормальных клеток в опухолевые (раковые). Эти вирусы вместо ДНК содержат РНК - их называют онкорнавирусами (сочетание из слов онкос - рак (греч.), неверно читаемого английского сокращения RNA (РНК) и слова вирус). Обратная транскриптаза вируса переводит его в форму ДНК, которая, включаясь в хромосому клетки, делает ее способной к неконтролируемому, злокачественному росту. Обратная транскриптаза стала одним из основных инструментов генных инженеров.
От РНК к белку. Генетический код. Вернемся к синтезу белка. Мы видим, что не сами гены (ДНК), а их РНК-копии служат матрицами для синтеза белковых молекул. Такие РНК называют информационными (иРНК) или матричными (мРНК). Их можно сравнить с рабочими чертежами (синьками), которые поступают в цеха завода (в нашем случае в цитоплазму), тогда как основные чертежи (ДНК) остаются в "конструкторском" бюро - ядре. По рабочим, матрицированным чертежам синтезируются белки.
Белки тоже полимерные макромолекулы: они состоят из полипептидных цепей, образованных остатками аминокислот. Вообще аминокислотой можно назвать любое соединение, содержащее одновременно аминогруппу (NH2) И группировку органической кислоты - карбоксильную (СООН). Число возможных аминокислот не очень отличается от бесконечности, но белки образуют только 20. Белковую цепочку называют полипептидной, потому что аминокислотные остатки в ней связаны пептидными связями (рис. 9): аминогруппа присоединяется к карбоксильной, при этом отщепляется молекула воды.
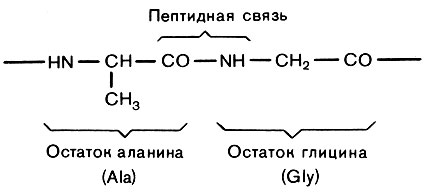
Рис. 9. Фрагмент полипептидной цепи
Ясно, что на одном конце полипептидной цепочки остается свободная аминогруппа, а на другом - карбоксильная. Эти концы и называют N-конец и С-конец. Все разнообразие белковых молекул в природе создается различием в длинах их полипептидных цепей и, главное, различным чередованием аминокислотных остатков в этих цепях.
Чередование аминокислотных остатков, или, иными словами, линейная последовательность расположения аминокислотных остатков в белковой молекуле, запрограммировано в нуклеотидной последовательности его гена (ДНК). Код, с помощью которого осуществляется такое программирование, получил название аминокислотного генетического кода. Мы часто будем встречаться с термином кодон, под которым понимают нуклеотидную комбинацию (т. е. комбинацию из A, G, Т и С), кодирующую один данный аминокислотный остаток.
Для того чтобы расшифровать аминокислотный генетический код, потребовалось более 10 лет упорнейшего труда целого научного коллектива. Одно наиболее общее свойство этого кода можем вывести и мы с вами. Известно, что в ДНК имеется четыре вида оснований - A, G, С и Т (в ее РНК-копии тоже четыре - A, G, С и U). Белки же построены из 20 видов аминокислот. Если бы код был однобуквенным, то мы бы смогли закодировать только 4 аминокислоты из 20. Двухбуквенного кода (т. е. с кодонами типа GA, AG, АА, CU и т. д.) также недостаточно, так как с его помощью можно закодировать только 42=16 аминокислот из 20. А вот трехбуквенным кодом (43=64) сочетания из А, Т, G и С можно закодировать все 20 аминокислот с избытком.
Трехбуквенность кода была доказана и в лаборатории английского ученого Ф. Крика. В тех же опытах было обнаружено, что два кодона, кодирующие соседние аминокислотные остатки, располагаются в полинуклеотидной цепи друг за другом, причем между ними нет вставок и они не перекрываются. Такой код называют неперекрывающимся кодом без запятых.

Генетический код оказался вырожденным, т. е. один и тот же аминокислотный остаток мог кодироваться разными кодонами. Нечто похожее было в старой русской азбуке: там тоже были буквы, читавшиеся одинаково ("е" и "ь", "i" и "и", "ф" и "θ "). Наш код вырожден неравномерно: аргинин, например, кодируется шестью кодонами, а метионин только одним.
В 1966 г. совместными усилиями ученых многих стран была завершена расшифровка генетического кода. Вот таблица, в которой даны кодоны для иРНК (рис 10).
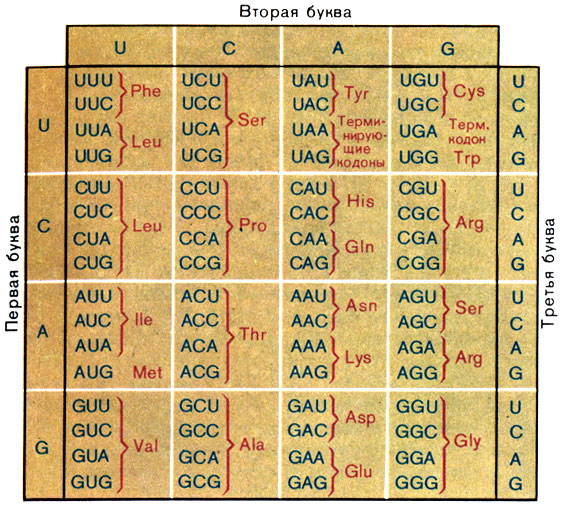
Рис. 10. Таблица аминокислотного генетического кода (кодовый словарь) Phe - фенилаланин; Leu - лейцин; Ile - изолейцин; Met - метионин; Val - валин; Ser - серин; Pro - пролин; Thr - треонин; Ala - аланин; Туr - тирозин; His - гистидин; Gin - глутамин; Asn - аспарагин; Lys - лизин; Asp - аспарагиновая кислота; Glu - глутаминовая кислота; Cys - цистеин; Тrр - триптофан; Arg - аргинин; Gly - глицин
Из нее видно, что 20 аминокислотам соответствует 61 кодон. Один, реже два из оставшихся трех кодонов стоят в конце кодирующей последовательности. Они соответствуют пропускам между словами в тексте.
Генетический код оказался универсальным для всей живой природы: бактерия, тигр и гвоздика используют одинаковые кодоны для соответствующих аминокислотных остатков. Лишь в особых внутриклеточных структурах - митохондриях, снабжающих клетки эукариот энергией, найдены незначительные отклонения от общего кода. Это свойство кода имеет огромное практическое значение: из него следует, что гены одного организма могут быть перенесены в любой другой организм (а не только из одной бактерии в другую, как это сделал Гриффите) и в любой чужеродной клетке содержащаяся в них программа может быть использована для синтеза полноценных белковых молекул.
Ф. Крик первым подметил, что между кодонами в РНК и аминокислотами не наблюдается никакого соответствия или химического сродства, и сделал отсюда вывод о существовании какой-то молекулы-посредника, "узнающей" одновременно и тройку нуклеотидов, и кодируемую ими аминокислоту. Эта догадка блестяще подтвердилась: посредниками оказались также молекулы РНК небольшой длины - транспортные РНК (тРНК). На одном из концов тРНК имеется последовательность из трех нуклеотидов, комплементарная кодону (ее называют антикодоном). А на другой присоединяется соответствующая этому кодону аминокислота (рис. 11). Связь кодон - антикодон временная, и в ней могут образовываться не только стандартные, канонические пары А-U и G-С, но и неканонические, менее прочные.
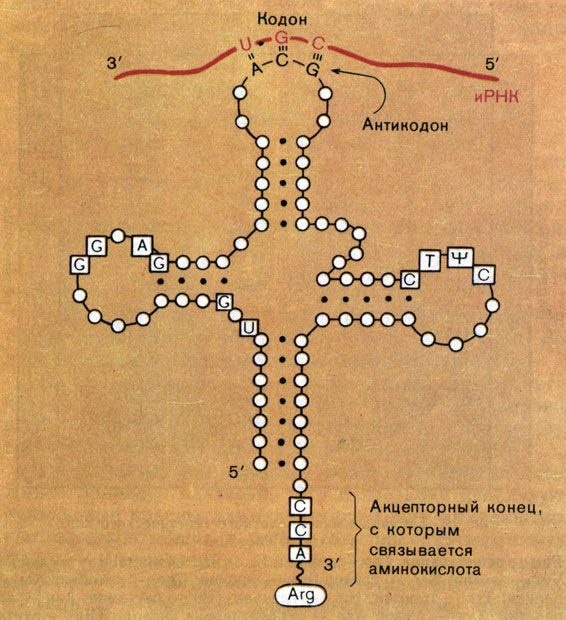
Рис. 11. Схема структуры транспортной РНК и кодонантикодоновых взаимодействий. Нуклеотидные остатки в тРНК изображены кружками. В квадратах находятся те нуклеотидные остатки, которые всегда присутствуют в тРНК в этих положениях. (Обратите внимание на то, что кодон в иРНК нужно 'читать' справа налево, так как 5'-конец РНК находится справа.)
С иРНК связывается, таким образом, не тРНК, а комплекс ее с соответствующей аминокислотой. Образование этих комплексов катализируется специальными ферментами - аминоацил-тРНК-синтетазами.
Легко догадаться, что каждая аминокислота должна иметь, по крайней мере, одну соответствующую ей тРНК. А так как код вырожденный, "сортов" тРНК гораздо больше 20.
Обратите внимание на еще одну тонкость. Где начало и где конец полипептидной цепи в белке? Установлено, что полипептиды в цитоплазме клеток синтезируются от N-конца к С-концу. N-конец договорились считать начальным. А он соответствует 5′-концу иРНК. Следовательно, 5′-конец иРНК начальный. Но ведь транскрипция, как и редупликация гена, идет так, что матричная нить прочитывается в направлении 3′-5′. Получается, что генетическая информация "перепечатывается" с конца, а читается, как водится, с начала!
Чтобы завершить эту картину, нам остается только добавить, что встреча тРНК, несущей на себе свою аминокислоту, с иРНК происходит на рибосоме. Рибосома, в свою очередь, построена из белков и РНК. Это уже третий класс РНК (рибосомные РНК).
События, происходящие на рибосоме, проще всего изобразить на схеме. Обратите внимание, что после образования каждой пептидной связи и ухода освободившейся тРНК иРНК перемещается на один кодон.
Рибосома начинает "читать" иРНК (молекулярные биологи называют синтез белка на рибосоме трансляцией) со строго определенного кодона (обычно им служит AUG), а заканчивает, как мы уже говорили, на терминирующем кодоне (рис. 12).
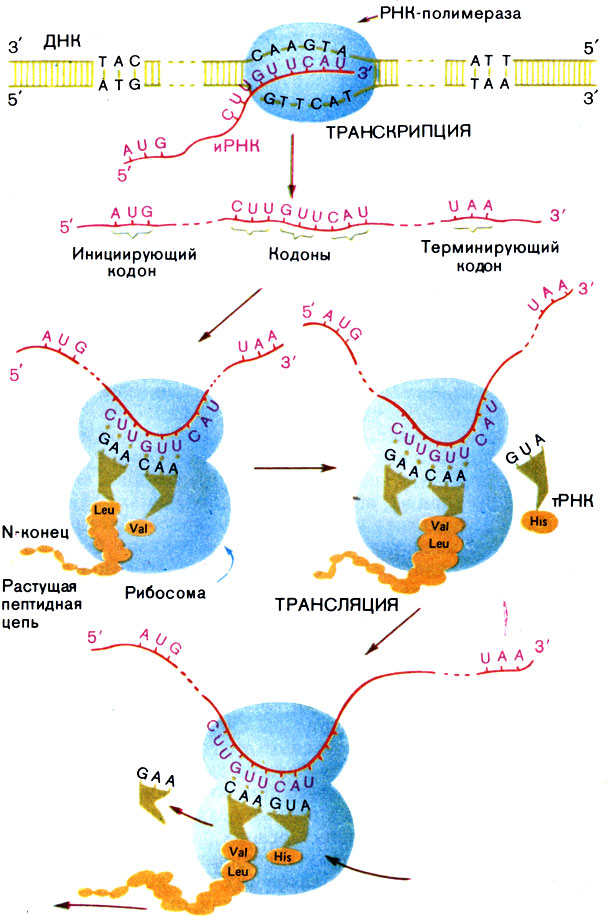
Рис. 12. Схема транскрипции (синтеза РНК на и трансляции (синтеза белка на рибосоме)
Многое из того, о чем мы сейчас рассказали, вы уже знаете из школьных учебников химии и биологии. Сейчас же вы познакомитесь с понятиями, более сложными и по своей новизне в учебники еще не попавшими. Постарайтесь запомнить все новые термины, которые вам попадутся. Не зная их, вы не поймете следующих глав.
Клетка управляет генами. Вскоре после разгадки генетического кода ученые поняли, что сделано было самое легкое, осталось, как это всегда бывает, самое трудное. В самом деле, что "запускает" в клетке одни гены и "выключает" работу других? Ген, кодирующий пищеварительный белок - пепсин, задействован только в специализированных клетках слизистой оболочки желудка. Что было бы, если бы он включился, начал транскрибироваться, например, в клетках нервной системы? Человек попросту переварил бы свой мозг. В конечном счете одна клетка отличается от другой, один орган развивается, отличаясь от другого, и один вид организмов отличается от другого только тем, что у них транскрипция идет с разных генов и разные белки синтезируются в разных количествах. Значит, основа развития организма даже не те гены, в которых закодированы аминокислотные последовательности белков (их называют структурными). Должна существовать какая-то система регуляции, включающая в определенное время одни гены и выключающая другие.
Такая система также входит в состав генома, какие-то последовательности ДНК должны быть регуляторными. Клетки млекопитающих способны синтезировать около 50 тыс. белков, а ДНК в клетке хватает на 5 млн. Получается, что генетическая система клеток в высшей степени "бюрократична": руководителей, т. е. регуляторных генов, там в тысячи раз больше, чем непосредственных исполнителей - структурных генов.
Иначе и быть не может. Человек при соответствующей подготовке может прочесть и понять любую книгу, написанную другим человеком: ведь его мозг - система несравненно более сложная, чем отдельная книга. А генетическая программа клетки должна сама себя прочесть, понять и задать клетке определенный объем работ.
Только сейчас ученые начинают постигать хитроумнейшие детали этого механизма. Чтобы лучше понять его, привлечем аналогии и наглядные примеры. Представим, что мы получили по радио серию букв и цифр, соответствующих названию судна и географическим координатам его положения. Ну и что? Смысл сообщения остается неясен. Но если сообщению предшествуют серия из 12 тире плюс несколько раз повторенное сочетание - три точки, три тире, три точки (SOS), оно расшифровывается однозначно - это сигнал бедствия. Если терпящий бедствие призывает помощь по радиотелефону, он должен перед этим произнести несколько раз условное легкоразличимое на слух буквосочетание МЭЙДЭЙ ("майский день" по-английски). Вот пример "регуляторной последовательности" - не из нуклеотидов, как в ДНК, а из точек и тире или звуков.
Рассуждая так, придем к выводу, что при каждом структурном гене должны быть последовательности, определяющие, когда, при каких условиях он должен быть транскрибирован, сколько актов транскрипции нужно сделать и когда он должен замолчать. Мало того, эти гены-регуляторы должны быть как-то связаны друг с другом. Только в этом случае клетки одного типа будут синтезировать один набор белков, клетки другого - другой и одна превратится, например, в эритроцит, другая в нейрон - клетку нервной системы.
Теперь посмотрим, что известно на сегодня о регуляторных элементах генов и генах-регуляторах.
Больше всего удалось узнать о регуляторных участках ДНК, расположенных непосредственно перед нуклеотидными последовательностями, кодирующими ту или иную полипептидную цепь белка. У них две обязанности. Во-первых, здесь находится последовательность-сигнал, показывающая, где необходимо связаться РНК-полимеразе, чтобы начать транскрипцию данного гена. Ее называют промотором. Во-вторых, в этом же участке ДНК расположены нуклеотидные последовательности, которые участвуют в регуляции уровня транскрипции: с одними из них связываются белки, усиливающие синтез иРНК на ДНК (белки-активаторы), а с другими, называемыми операторами, белки-репрессоры, которые подавляют транскрипцию или вовсе ее останавливают. Ясно, что белки-активаторы и белки-репрессоры также должны быть закодированы в каких-то генах. Такими генами являются гены-регуляторы.
Чтобы нагляднее представить механизм действия, рассмотрим, как регулируется работа гена, в котором закодирован фермент β-галактозидаза у бактерии - кишечной палочки. Если бактерия синтезирует этот фермент, она может расти, потребляя в качестве единственного источника углерода углевод молочный сахар - лактозу. Обычный источник углерода для кишечной палочки - глюкоза. Лактоза - это дисахарид, она содержит остатки галактозы и глюкозы. Если вместо глюкозы "кормить" кишечную палочку лактозой, то в ней начинается синтез фермента β-галактозидазы, выщепляющей из лактозы необходимую бактерии глюкозу. Обратите внимание, если глюкоза присутствует в питательной среде, то кишечная палочка β-галактозидазы не образует. Она ей просто не нужна. На языке генетики это означает, что ген β-галактозидазы репрессирован, т. е. "молчит". Но при замене глюкозы на лактозу он включается.
Впервые расшифровать события, которые здесь происходят, сумели французские ученые Ж. Моно и Ф. Жакоб. Они установили, что, когда кишечная палочка растет на глюкозе, промотор гена β-галактозидазы недоступен для РНК-полимеразы потому, что он заблокирован белком-репрессором, прочно связанным с оператором (рис. 13). Репрессор можно сравнить с пломбой на двери запечатанного помещения: эта пломба срывается в каких-то чрезвычайных обстоятельствах.
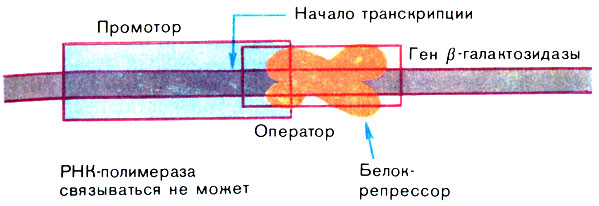
Рис. 13. Регуляция работы гена (ген β-галактозидазы репрессирован)
Заменим глюкозу на лактозу. Промотор гена β-галактозидазы свободен, и синтез иРНК на этом гене может начаться, так как репрессор теперь находится в неактивном состоянии. Что же с ним произошло? Оказывается, новый источник углерода (равно как и многие другие соединения, похожие на лактозу) прочно связался с репрессором и сделал его неспособным взаимодействовать с оператором. Такие вещества называют индукторами. Более того, в отсутствие глюкозы в клетке начинает работать белок-активатор, который связывается с ДНК слева от РНК-полимеразы и помогает ей начать транскрипцию (рис. 14).
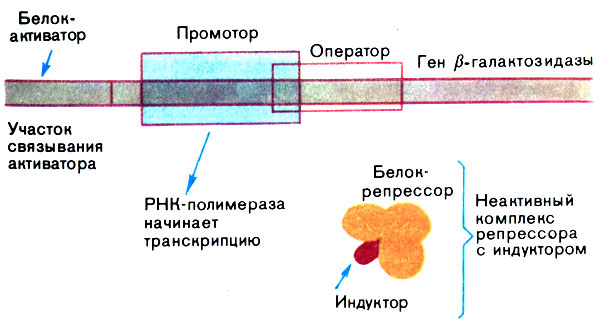
Рис. 14. Регуляция работы гена (ген β-галактозидазы активен)
Такое состояние будет поддерживаться до тех пор, пока в среде есть индуктор. Любопытно, что кишечную палочку можно "обмануть": подсунуть ей соединение, которое связывается с репрессором, но не расщепляется белком - продуктом репрессированного гена. Ген тогда работает вхолостую.
Схема Жакоба и Моно - один из самых простых способов регуляции генной активности, но далеко не единственный. Например, включать одни и выключать другие гены можно, изменяя саму РНК-полимеразу. Этот фермент состоит из многих полипептидных цепей, называемых субъединицами. Присутствие одной из таких субъединиц (называемой δ-фактором) абсолютно необходимо для того, чтобы РНК-полимераза связалась с промотором и начала транскрипцию. Изменяя δ-фактор, можно изменять сродство РНК-полимеразы к промоторам. Таким путем регулируются гены во время так называемого "теплового шока" - явления, широко распространенного в природе. Если любой живой организм (будь то бактерия или мушка-дрозофила) поместить в условия повышенной температуры (не смертельной, конечно, но при которой он себя чувствует неуютно), то организм будет приспосабливаться. Он в срочном порядке начинает в больших количествах синтезировать спасательные белки (или белки-спасатели). Уровень синтеза обычных белков при этом сильно снижается. В этом случае активизируются одни гены (гены "теплового шока") и подавляются другие гены. Оказалось, что, по крайней мере, у бактерий активация генов "теплового шока" связана с заменой старого δ-фактора на новый, приспособленный к промоторам этих генов.
Говоря о регуляторных элементах генов, нельзя не упомянуть еще о нескольких. Во-первых, недавно в хромосомах были найдены участки - усилители транскрипции, которые расположены на большом (иногда очень большом) расстоянии от кодирующей последовательности гена и его промотора. Механизм их действия - одна из самых интересных загадок молекулярной биологии. Во-вторых, для нормальной работы гена очень важны участки, расположенные в конце кодирующей последовательности и служащие сигналом остановки транскрипции. Их называют терминаторами. В некоторых генах можно обнаружить терминатор и внутри кодирующей последовательности. Такой ген может полностью транскрибироваться только в присутствии так называемого белка-антитерминатора, позволяющего РНК-полимеразе "проскочить" внутренний терминатор.
Теперь можно воссоздать достаточно точную картину расположения генетических регуляторных элементов, известных в настоящее время (рис. 15). Не следует забывать, однако, что многое и в организации генов, и в их регуляции еще предстоит выяснить в будущем, и эта схема еще будет существенно дополнена. Молекулярная биология очень молодая наука, и еще много открытий впереди. Посмотрите, например, как постоянно изменяется само понятие гена.
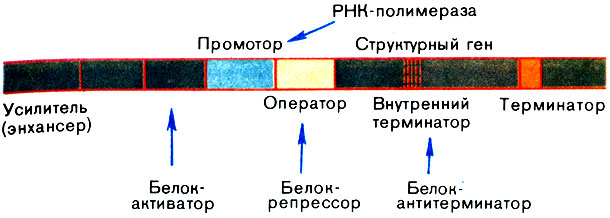
Рис. 15. Регуляторные элементы гена
Так что же такое "ген"? Как мы уже рассказывали, классическая генетика пришла к представлению "один ген - один белок". Когда стало ясно, что гены построены из ДНК, появилось определение: ген - это линейный участок ДНК, в котором закодирован один белок.
Затем стало известно, что многие белки (в частности, РНК-полимераза) построены из нескольких полипептидных цепей (субъединиц). Пришлось опять модифицировать определение гена: ген - это линейный участок ДНК, в котором закодирована одна полипептидная цепь белка. Однако около 10 лет назад выяснилось, что и это определение следует изменить. Было установлено, что кодирующие последовательности многих генов эукариот содержат вставки, которые к данному белку отношения не имеют. Эта особенность организации генов настолько важна и настолько широко распространена, что следует более подробно рассказать об этом. Такое открытие стало возможным, когда в руках ученых оказались, с одной стороны, индивидуальные гены, а с другой - индивидуальные иРНК для определенных белков. (В последующих главах мы расскажем, как этого можно достичь.) Вы знаете, что иРНК - это комплементарная копия гена. Каково же было удивление ученых, когда, получив ДНК-РНК-гибрид из денатурированной ДНК некоторых генов и транскриптов с них (иРНК), они увидели в электронный микроскоп такую картину (рис. 16).
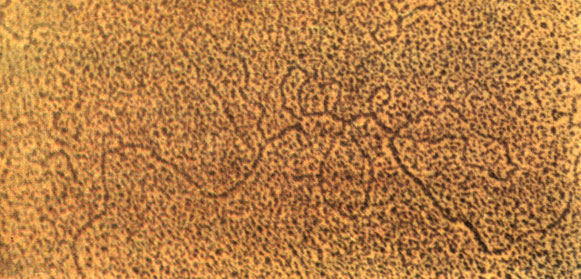
Рис. 16. Гибридизация гена белка овальбумина с и РНК этого белка. На микрофотографии показано, как гибрид (ДНК-РНК) выглядит под электронным микроскопом. Ниже приведина схема, объясняющая эту микрофотографию
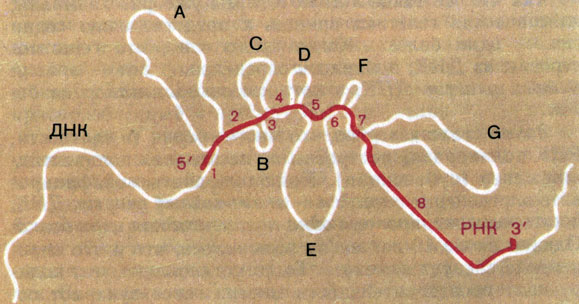
Протяженные участки ДНК (здесь они выглядят как петли) вообще не гибридизовались с РНК! Но ведь именно эта иРНК транслировала и давала полноценный белок. Более того, ее выделили из комплекса с рибосомами во время трансляции. Здесь вспомнили, что транслируемая иРНК обычно образуется из более длинного предшественника - пре-иРНК. Если с геном сгибридизировать такой предшественник, то гибрид выглядит нормально (рис. 17).

Рис. 17. Гибрид гена овальбумина с предшественником иРНК
Значит, в процессе превращения пре-иРНК в иРНК из его цепи исчезли какие-то внутренние участки. Это можно сделать, только разрезав цепь в определенных местах, удалив эти участки и вновь сшив оставшиеся районы РНК, совокупность которых кодирует данную полипептидную цепь (рис. 18). Этот процесс называют сплайсингом (от английского слова, попавшего в русский язык через морской жаргон: "сплеснивать" конец значит сращивать канат, веревку из отдельных кусков). Ведь когда внутренние участки вырезаны, остающиеся должны быть каким-то образом соединены в единую последовательность.
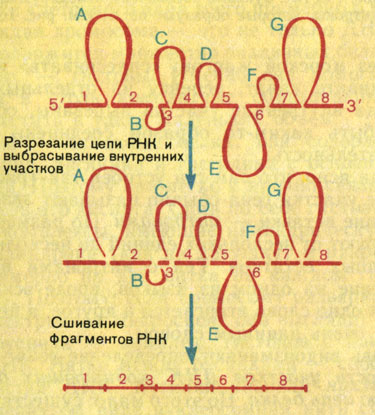
Рис. 18. Схема сплайсинга иРНК гена овальбумина. Цифрами обозначены кодирующие последовательности - экзоны, буквами - некодирующие вставки - интроны
Отсюда ясно, что сам ген устроен так (рис. 19). Кодирующие участки гена обычно называют экзонами, некодирующие вставки - интронами. По размерам интроны варьируют от нескольких единиц до нескольких тысяч нуклеотидных остатков. Ген с интронами напоминает предложение на одном из языков, вроде эскимосского, в которых одно слово вторгается в другое, а целая фраза выглядит очень длинным словом.
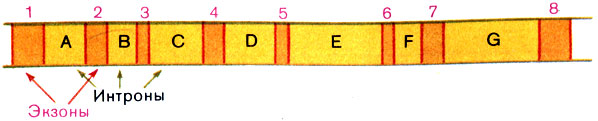
Рис. 19. Схема строения гена овальбумина. Цифрами обозначены кодирующие последовательности - экзоны, гибридизирующие с иРНК. Буквами обозначены некодирующие вставки - интроны, которые образуют петли (см. рис. 16)
И вновь видоизменяют определение гена: ген - это совокупность участков ДНК, кодирующих одну полипептидную цепь белка. Но этого мало: существуют и гены транспортных и рибосомных РНК, не кодирующие белков. А последовательности-усилители - гены это или нет? Ведь мутация не только в гене, кодирующем белке, но и в усилителе или промоторе, интроне или терминаторе может изменять, как это показано, общий облик и физиологию организма, его фенотип. Это изменение передается по наследству, а ведь наследственность, как мы знаем, - функция генов. Может быть, правильнее такая обтекаемая формулировка: ген - нуклеотидная последовательность, выполняющая определенную функцию?
Каким же образом гигантские по длине полинуклео- тидные цепи ДНК организованы в клеточном ядре?
Главная черта этой организации - необычайная компактность ДНК. Рассмотрим простейшую систему - ДНК-содержащий вирус, например аденовирус, который вызывает ОРЗ (острое респираторное заболевание, или обычную простуду). Если частицу этого вируса рассматривать как шарик, т. е. диаметр составляет всего 0,07 мкм, ДНК же аденовируса имеет длину 11 мкм.
Еще более удивительным выглядит сравнение размеров клеточного ядра клетки человека и его ДНК. Если мысленно ДНК, содержащуюся во всех 46 хромосомах человеческой клетки, соединить в одну нить, то ее длина будет равна 1,8 м! (Не забывайте, что диаметр такой нити всего 20 десятимиллиардных долей метра; представьте провод от Земли до Луны диаметром в 4 мм.) Диаметр же клеточного ядра всего примерно 6 мкм.
Но каждая хромосома - это не только ДНК. В хромосомах содержится множество различных белков. Главные из них - гистоны. Именно они помогают ДНК сложиться в столь компактные образования, что их можно уместить в такой маленький объем.
Гистоны - основные белки: они содержат очень много лизина, аргинина и гистидина с основными аминогруппами. Поэтому они легко и прочно связываются с ДНК - ведь это кислота. Сами по себе гистоны объединяются в комплексы, имеющие цилиндрическую форму. На эти цилиндрики (как на катушку) и накручивается ДНК (примерно по два витка на каждом гистоновом комплексе). Поэтому, если хромосому развернуть в нитку, то в электронном микроскопе она выглядит как бусы. Каждая бусинка - комплекс гистонов с накрученной на него ДНК. Нитки между бусинками - свободные от белка участки ДНК. Бусинки назвали нуклеосомами.
В развернутом виде нуклеосомная нить существует только в искусственных условиях. В хромосоме отдельные нуклеосомы объединяются по шесть или восемь штук и образуют тяжи, которые, в свою очередь, перекручены друг с другом в более толстые тяжи, и так до тех пор, пока не возникнет огромная хромосомная структура.
Вот таким образом и удается спрятать длинную ДНК в очень малый объем. Правда, упакованные таким образом гены неактивны. РНК-полимераза, к примеру, к ним просто не может пробраться. Поэтому перед началом работы генов целая область хромосомы, в которой они находятся, раскручивается и переходит на более низкий уровень организации. В этом процессе заключен еще один способ регуляции активности генов: в клетке могут включаться и выключаться целые группы генов в зависимости от того, в какого типа хромосомной структуре они находятся.
Мы дали очень упрощенную картину сегодняшних знаний о структуре гена - ровно столько, чтобы можно было понять принципы генной инженерии. Но уже из нее становится ясно, в какое сложное устройство приходится вторгаться, чтобы направленно изменить наследственность. Ген, как смерть Кощея Бессмертного, запрятан в чрезвычайно сложной структуре за оболочкой ядра; очень непросто его оттуда извлечь, отделить от других генов и вставить в другой геном, так чтобы он работал в новом месте.
Выделять ДНК в чистом виде научились давно. Но в растворе эти длиннейшие молекулы представляют собой хаотическую смесь обломков, которую тепловое движение все время перебалтывает.
Представьте содержание большой книги (например, однотомника собрания сочинений Пушкина), напечатанное точками и тире на телеграфной ленте. Тираж, скажем 1 млн. экземпляров, собран в стог, который непрерывно ворошат. Телеграфные ленты при этом рвутся в случайных местах. Требуется дистанционно с большого расстояния взять оттуда ленту, на которой отпечатано стихотворение "Анчар" или поэма "Полтава" (примерно такое соотношение между длинами самого маленького и самого большого гена).
Перед будущими генными инженерами встала задача огромной сложности: ведь первичные структуры, т. е. последовательности нуклеотидов в генах, были практически неизвестны. Зная генетический код, можно было приблизительно, с учетом вырожденности кода, прогнозировать последовательности структурных генов, тех самых, которые кодируют белки. Однако узнать последовательность нуклеотидов в промоторах или усилителях стало возможным лишь тогда, когда их научились выделять. Образно говоря, уже проделав операцию по извлечению нужной телеграфной ленты из целого стога, не только не видя ее, но и не зная азбуки Морзе!
Еще недавно такая задача казалась, если и разрешимой, то в очень отдаленном будущем, и это заманчивое будущее отодвигалось с каждым новым открытием. Похоже, подтверждалась древняя грустная мудрость: "Во многом знании много печали". Но внезапно наступил перелом. Помощь пришла из, казалось бы, узкой специальной отрасли - генетики фагов - вирусов микроорганизмов и генетики самих бактерий.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'