XIV. Отношение 9:3:3:1. Отношение 1:1:1:1
Теперь мы знаем все необходимое, чтобы понять результаты скрещиваний между организмами, различающимися более чем по одному менделевскому признаку. (Под менделевским признаком подразумевается признак, определяемый аллелями одного гена.) Сформулируем еще раз правила, которые необходимо помнить.
1. В формулах генотипов родителей каждый признак должен быть обозначен парой генетических символов. (В качестве генетического символа используют букву или знак +). Например, скрещивание мыши альбиноса (♀cc) с самцом, имеющим волнистую шерсть (♂ ww), можно записать следующим образом:
Это скрещивание нельзя записывать как ♀ cc × ♂ ww, поскольку такая форма записи часто приводит к ошибкам.
2. Каждая гамета несет по одному аллелю каждой пары генов. При скрещивании организмов с признаками, определяемыми шестью парами генов, каждая гамета несет шесть генов. Исключение составляют гены, сцепленные с полом, которые не имеют аллелей у организмов гетерогаметного пола. Например, все сперматозоиды, которые образует мышь-альбинос с волнистой шерстью (cc ww), имеют генотип cw. Наоборот, голубоглазый мужчина, страдающий дальтонизмом (bb cb), образует два типа гамет: сперматозоиды с Х-хромосомой имеют генотип b сb, а сперматозоиды с Y-хромосомой - генотип b.
3. Если ген представлен парой одинаковых аллелей, то этот аллель находится во всех гаметах. Если ген представлен парой разных аллелей, то половина гамет несет один аллель, а половина - другой (первый закон Менделя).
4. Если несколько пар генов представлены разными аллелями, то гаметы, имеющие все возможные комбинации аллелей, будут встречаться с равной частотой (второй закон Менделя).
Пользуясь этими правилами, можно предсказать результаты скрещивания между организмами с признаками, определяемыми двумя парами аллелей, например между красноцветковыми растениями гороха с ребристыми бобами и белоцветковыми растениями с гладкими бобами (рис. 34). Аллели, определяющие красную и белую окраску цветков, обозначены соответственно буквами R и r, а аллели, определяющие гладкую и ребристую форму бобов, - буквами Р и р. Красная окраска цветков доминирует над белой, а гладкая форма бобов - над ребристой.
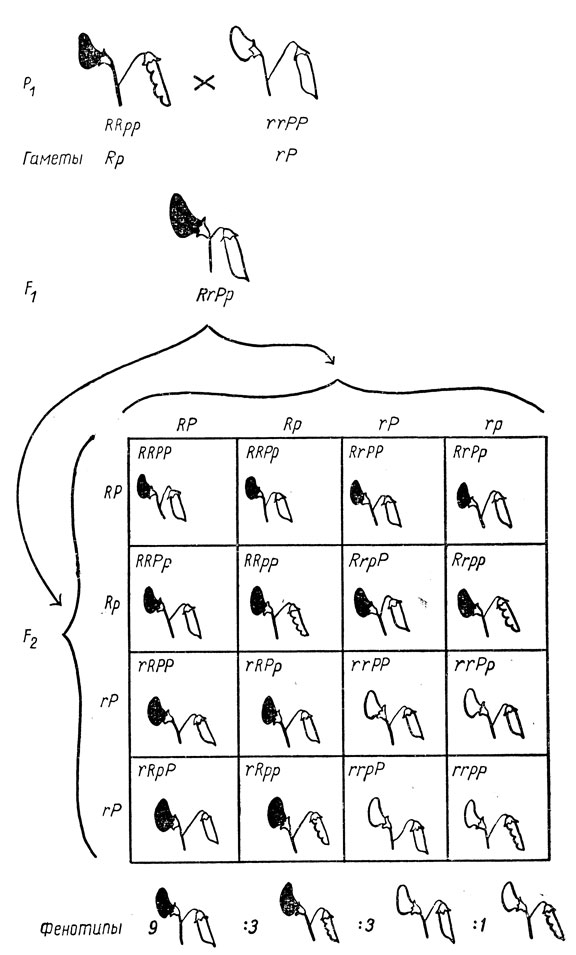
Рис. 34. Скрещивание растений гороха, отличающихся друг от друга по двум парам несцепленных генов: R - ген, определяющий красную окраску цветков; r - его аллель, определяющий белую окраску цветков; Р - ген, определяющий гладкую форму бобов; р - его аллель, определяющий ребристую форму бобов
Родительские растения гомозиготны по обеим парам генов, и поэтому каждое из них образует только один тип гамет. Растения F1 являются гетерозиготными по обеим парам генов и образуют четыре типа гамет - те самые четыре типа, которые были изображены на рис. 31. Поскольку как мужские, так и женские гаметы представлены этими четырьмя типами, решетка, характеризующая растения F2, состоит из шестнадцати (4 × 4) квадратов.
Вы видите в некоторых квадратах решетки растения с одинаковыми фенотипами, которые суммированы в нижней части рисунка. Нетрудно заметить, что в F2 помимо красноцветковых растений с ребристыми бобами и бело-цветковых с гладкими бобами имеются два новых типа: красноцветковые растения с гладкими бобами и бело-цветковые с ребристыми бобами. Эти четыре фенотипа растений F2 представляют все возможные сочетания двух пар признаков. Таким образом, свободное сочетание несцепленных генов приводит не только к образованию всех возможных сочетаний этих генов в гаметах, но и к появлению всех возможных сочетаний соответствующих признаков в фенотипах. Однако есть одно существенное различие: в гаметах все комбинации генов встречаются с одинаковой частотой, в то время как в фенотипах частота сочетаний разных признаков может быть различной. В данном случае отношение фенотипов оказывается следующим: 9 растений с обоими доминантными признаками : 3 растения с доминантной окраской цветков и с рецессивной формой бобов : 3 растения с рецессивной окраской цветков и с доминантной формой бобов : 1 растение с обоими рецессивными признаками. Частота этих фенотипов не зависит от сочетаний признаков у родителей. Например, скрещивание между красноцветковым растением с гладкими бобами (RR РР) и белоцветковым растением с ребристыми бобами (rr рр) даст такие же растения (Rr Рр) и, следовательно, то же отношение фенотипов в F2 (9:3:3:1).
Если вы вернетесь теперь к эксперименту Менделя, описанному в начале главы XIII, то увидите, что отношение 315:101:108:32 действительно очень близко к отношению 9:3:3:1. Именно на основании анализа этого отношения Мендель и сформулировал свой второй закон. Отношение 9:3:3:1 характерно для F2, полученного в результате скрещивания между организмами (Р1), различающимися двумя парами аллелей.
Почему же отношение числа гамет разных типов, равное 1:1:1:1 у растений F1, приводит к отношению фенотипов потомства F2, равному 9:3:3:1? При рассмотрении решетки (см. рис. 34) становится ясным, что это обусловлено доминированием красной окраски цветков над белой и гладкой формы бобов над ребристой. Для того чтобы узнать, какие гаметы образуются растениями с одинаковыми фенотипами в F2, необходимо избежать осложнений, создаваемых доминированием. Это можно сделать с помощью так называемого возвратного скрещивания, что и проиллюстрировано на рис. 35 на примере скрещивания двойной гетерозиготы с двойным рецессивом.
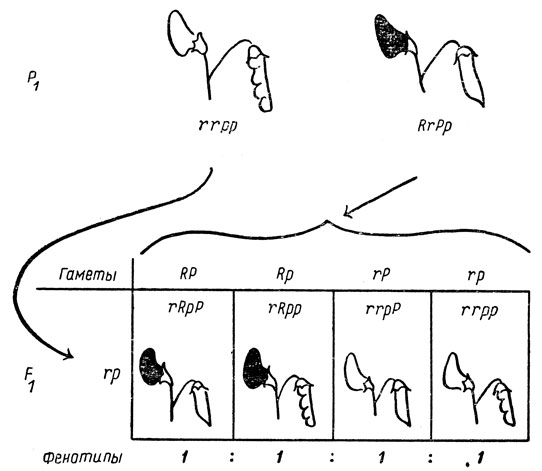
Рис. 35. Возвратное скрещивание двойной гетерозиготы (Rr Рр) с двойным рецессивом (rr рр)
Растение с генотипом Rr Рр образует гаметы четырех типов: RP, Rp, rP и rр, а растение с генотипом rr рр формирует гаметы только одного типа - гр. Поэтому решетка состоит только из четырех квадратов. Это по существу то же анализирующее скрещивание, которое было описано в главе VI. Действительно, его можно использовать, чтобы установить, не является ли красно-цветковое растение с гладкими бобами гетерозиготным по окраске цветков, или по форме бобов, или одновременно по двум этим признакам. В табл. 14 показаны возможные результаты такого скрещивания.

Таблица 14
В общем, независимо от числа рассматриваемых пар генов скрещивание особи с организмом, гомозиготным по всем рецессивным аллелям, можно использовать как анализирующее скрещивание. Оно всегда дает отношение фенотипов 1:1:1.
Второй закон Менделя, как и первый, является статистическим. Поэтому отношения, которые он предсказывает, будут строго выполняться только при очень большом числе потомков. Если бы Мендель имел всего 16 растений F2, он вряд ли получил бы отношение их фенотипов точно 9:3:3:1. В частности, он мог бы при этом вообще не получить двойного рецессива, который представлен только 1/16 растений F2. Это очень важно иметь в виду тем, кто хочет получить новый тип организма путем скрещиваний. Предположим, человек, разводящий кроликов, хочет вывести линию длинношерстных (ангорских) кроликов альбиносов из альбиносов и черной ангорской линии. Нам уже известно, что ген, вызывающий альбинизм (с), рецессивен Ген, определяющий длинную шерсть (l), также рецессивен. Поэтому первое скрещивание можно записать следующим образом:
Вы видите, что в соответствии с первым правилом в формулу генотипа кролика альбиноса введен ген L, который является аллелем гена l и определяет короткую шерсть, а в формулу генотипа ангорского кролика - ген, обозначенный буквой С, который является аллелем гена с и определяет черную окраску шерсти. (Гены, вызывающие окраску неагути (аа) у ангорского кролика, в формулу не введены как несущественные при данном скрещивании.)
Животные F1 имеют генотип Сс Ll. При скрещивании их между собой в F2 будут получены кролики четырех фенотипов в отношении 9:3:3:1. Если их число достаточно велико, то 9/16 кроликов проявят оба доминантных признака, т. е они не будут ни альбиносами, ни ангорскими, 3/16 кроликов будут иметь сочетание доминантного и рецессивного признаков (черные ангорские кролики), 3/16 кроликов будут иметь другое сочетание доминантного и рецессивного признаков (альбиносы с короткой шерстью) и, наконец, 1/16 кроликов проявят оба рецессивных признака, т. е. они будут альбиносами и ангорскими. Именно этот последний тип белых ангорских кроликов и интересовал селекционера. Если иметь в виду, что каждый помет состоит из 7-8 крольчат, то необходимо совсем немного пометов, чтобы отобрать самца и самку, которые дали бы начало нужной линии.
Поскольку в F2 кроликов с доминантными фенотипами больше, чем с рецессивными, можно было бы думать, что для выведения линии хотя бы с одним доминантным признаком достаточно меньшего Числа пометов. Однако это не так. Предположим, селекционер, имеющий линию ангорских альбиносов (сс ll), хотел бы иметь линию окрашенных ангорских кроликов (СС ll). Он скрещивает своих ангорских альбиносов с кроликами дикого типа (окрашенные, с короткой шерстью), имеющими генотип СС LL. Это можно записать так: сс ll × СС LL. F1 состоит из кроликов с генотипом СсЫ. Скрещивание их между собой дает следующее отношение кроликов в F2:9 окрашенных с короткой шерстью : 3 окрашенных ангорских : 3 альбиноса с короткой шерстью : 1 ангорский альбинос.
Действительно, в этом случае из 16 животных 3 кролика будут иметь желаемый селекционером фенотип (окрашенные ангорские). Но селекционеру нужны кролики этого фенотипа, только гомозиготные по соответствующим генам; в то же время из 3 окрашенных ангорских кроликов 2 оказываются гетерозиготными (Сс). Если это непонятно, вернитесь к растениям F2 (см. рис. 34), которые, подобно окрашенным ангорским кроликам, проявляют 1 доминантный и 1 рецессивный признак (3 красно-цветковых растения с ребристыми бобами или 3 белоцветковых растения с гладкими бобами). Действительно, в каждом случае 2 растения из 3 гетерозиготны по одному из двух генов и только 1 гомозиготно по обоим генам. Такие гомозиготные растения составляют 1/16 часть F2, так же как и двойные рецессивы - белоцветковые растения с ребристыми бобами или ангорские кролики-альбиносы.
Поэтому при выведении путем скрещиваний новых чистопородных линий как по доминантным, так и по рецессивным признакам необходимо одинаковое число животных. В действительности же легче получить линию ангорских альбиносов, чем ангорских окрашенных кроликов. Хотя последних в F2 больше, но они требуют испытания по потомству, в то время как все ангорские альбиносы являются, как известно, гомозиготными.
В рассмотренных выше скрещиваниях мы имели дело с парами аллелей, один из которых полностью доминировал над другим. В противном случае гетерозиготу по одному или обоим генам можно отличить от любой гомозиготы. Отсутствие полного доминирования даст в F2 дополнительные фенотипы.
Вы сами разберете соответствующий пример при выполнении заданий к этой главе.
Выводы
1. Свободное сочетание несцепленных генов в гаметах определяет свободное сочетание несцепленных признаков у потомства. Родители, отличающиеся друг от друга по нескольким несцепленным менделевским признакам, дадут все возможные сочетания этих признаков у детей или более отдаленных потомков.
2. Скрещивание между индивидуумами, различающимися двумя парами генов, дает в F1 организмы, у которых проявляются оба доминантных аллеля; в F2 организмы с четырьмя возможными сочетаниями признаков находятся в следующем соотношении:
а) 9 из 16 проявляют оба доминантных признака;
б) 3 из 16 проявляют доминантный признак первой пары генов и рецессивный признак второй пары;
в) 3 из 16 проявляют рецессивный признак первой пары генов и доминантный признак второй пары;
г) 1 из 16 проявляют оба рецессивных признака.
Только 1 из 9 организмов в пункте а) и 1 из 3 в пунктах б) и в) гомозиготны, т. е. чистопородны по этим признакам (см. рис. 34).
3. Отношение 9:3:3:1 не зависит от сочетаний доминантных и рецессивных признаков у родителей (будут ли оба доминантных признака у одного из родителей или один из них будет у матери, а другой - у отца).
4. Если гетерозиготу по нескольким генам, определяющим несцепленные менделевские признаки, скрестить с гомозиготой по тем же генам, определяющим рецессивные признаки, то среди особей F1 будут все возможные комбинации этих признаков в отношении 1:1:1.
5. Независимо от числа рассматриваемых пар генов для выявления рецессивных аллелей у гетерозиготного организма в качестве анализирующего скрещивания может быть использовано возвратное скрещивание с организмом, гомозиготным по соответствующим рецессивным аллелям.
Задание
1. У крупного рогатого скота ген комолости (Р) доминирует над его аллелем (р), определяющим образование рогов. Аллели, определяющие белую (W) и окрашенную (w) шерсть, не доминируют один над другим, так что гетерозиготные животные оказываются чалыми. В приложении 5 изображены животные F1, полученные в результате скрещивания между окрашенным рогатым быком и белой комолой коровой, гомозиготной по аллелям, определяющим оба признака. Скопируйте решетку и заполните ее. Получите окончательное отношение животных F2 с разными фенотипами. Число потомков от одного такого скрещивания слишком мало, чтобы получить ожидаемое отношение, но при достаточно большом числе животных это отношение несомненно получится.
2. Вернитесь к заданию 2 (глава XIII), где перечислены несцепленные гены мыши.
Предполагалось, что самец с окрашенной королевской шерстью, длинными ушами и скрученным хвостом гетерозиготен по одному или нескольким генам: альбинизм (с), гладкая шерсть (r), короткие уши (s) и прямой хвост (k). Чтобы это проверить, было проведено возвратное скрещивание этого самца с самками-альбиносами с гладкой шерстью, короткими ушами и прямым хвостом (сc rr ss kk). Потомство состояло из следующих животных:
- четыре животных с окрашенной королевской шерстью, длинными ушами, скрученным хвостом;
- три животных с окрашенной королевской шерстью, короткими ушами, скрученным хвостом;
- пять животных-альбиносов с королевской шерстью, длинными ушами, скрученным хвостом;
- четыре животных-альбиноса с королевской шерстью, короткими ушами, скрученным хвостом;
- два животных с окрашенной королевской шерстью, длинными ушами, прямым хвостом;
- три животных с окрашенной королевской шерстью, короткими ушами, прямым хвостом;
- пять животных-альбиносов с королевской шерстью, длинными ушами, прямым хвостом;
- четыре животных-альбиноса с королевской шерстью, короткими ушами, прямым хвостом.
Определите генотип самца.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'