II.2. Жизненные циклы, способы и особенности размножения у высших растений
Большая часть жизненного цикла у высших растений также представлена диплофазой - спорофитом, т. е. собственно растением; гаплоидная фаза ограничена гаметофитом и гаметами. У растений наблюдается большое разнообразие жизненных циклов в связи с наличием и чередованием у многих видов разных типов размножения: полового, бесполого и апомиксиса.
Половому размножению, также как и у животных, предшествует образование гамет, которое осуществляется в две стадии - спорогенеза и гаметогенеза.
В течение микроспорогенеза диплоидная археспориальная клетка - микроспороцит - претерпевает мейоз, в результате формируется четыре гаплоидных микроспоры. На стадии гаметогенеза каждая микроспора претерпевает два митотических деления. В результате первого образуются вегетативная и генеративная клетки. Вегетативная клетка и ее ядро больше не делится, в ней накапливаются питательные вещества, обеспечивающие прорастание пыльцевого зерна. Формируется двухклеточное пыльцевое зерно - начало развития мужского гаметофита (у бобовых). При прорастании на рыльце генеративная клетка вновь делится и образуются две мужские гаметы - спермии (рис. II. 3, а). У других видов, например у злаков, деление генеративной клетки осуществляется в пыльцевом зерне и формируется трехклеточное пыльцевое зерно, содержащее два спермия.
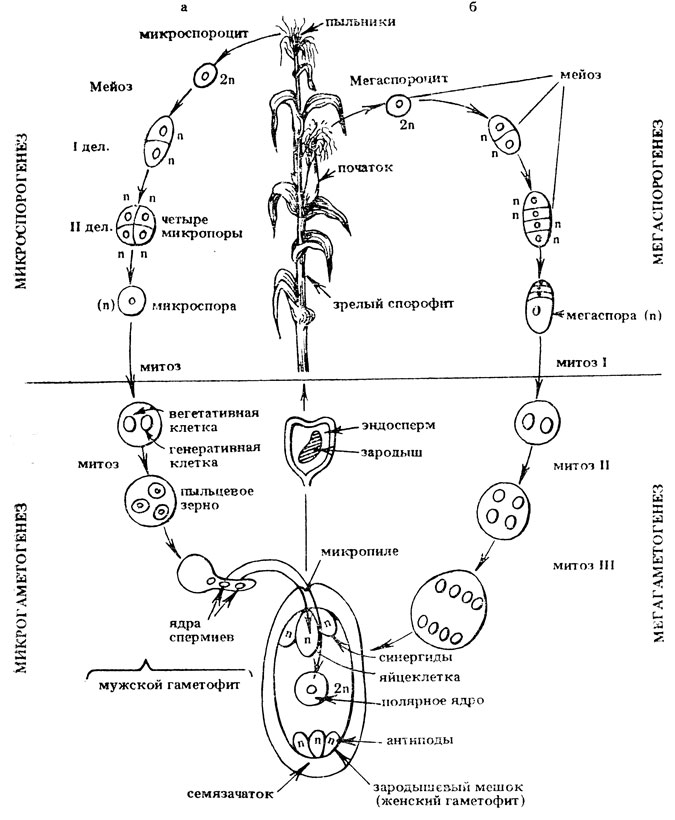
Рис. II.3. Жизненный цикл кукурузы: а - микроспорогенез и микрогаметогенез, б - мегаспорогенез и мегагаметогенез
При мегаспорогенезе (или макроспорогенезе) диплоидная археспориальная клетка растет, превращаясь в материнскую клетку мегаспоры (мегаспороцит), которая делится мейотически. Образуется 4 гаплоидных клетки, из которых лишь одна превращается в гаплоидную мегаспору, а остальные три - дегенерируют. В процессе мегагаметогенеза ядро гаплоидной мегаспоры трижды делится митотически, возникает 8 наследственно одинаковых ядер. Формируется женский гаметофит - зародышевый мешок, содержащий на одном полюсе яйцеклетку (женскую гамету) и две синергиды, на другом - три антиподы, в центре два полярных ядра, которые, сливаясь, образуют диплоидное центральное ядро или полярную клетку (рис. II. 3, б). Мегаспоры располагаются линейно. Зародышевый мешок может развиваться из любой мегаспоры, или, как, например, у пшеницы, кукурузы и др. он развивается из базальной мегаспоры, наиболее удаленной от микропиле. В некоторых случаях это может оказывать влияние на характер расщепления (см. гл. III). Рассмотренная схема образования восьми-ядерного зародышевого мешка из одной мегаспоры наиболее типична для высших растений. Однако у некоторых групп растений этот процесс может протекать по-другому.
Для полового процесса высших растений характерно наличие двойного оплодотворения. Оно заключается в том, что в зародышевый мешок проникает два спермия. Один из них сливается с ядром яйцеклетки, образуя диплоидную зиготу, из которой развивается зародыш, второй - с диплоидным полярным ядром, образуя триплоидный эндосперм. В одном пыльцевом зерне формируются одинаковые по генотипу спермин и в оплодотворении обычно участвуют спермии лишь одной пыльцевой трубки. Однако, за счет открытого цветения некоторых цветков или из-за недостаточно тщательной изоляции, на рыльце может прорасти чужеродная пыльца. Если материнское растение рецессивно по признакам эндосперма, а чужеродная пыльца несет доминантный аллель, то благодаря слиянию одного из спермиев с полярным ядром, имеющим рецессивный генотип, появляются гибридные семена, отличающиеся от семян, возникших в результате самоопыления: они проявляют доминантный признак. Например, при самоопылении растений кукурузы с неокрашенными семенами (рецессивный признак) на початках иногда обнаруживаются окрашенные семена (доминантный признак), что указывает на гибридную природу этих семян. Это явление - проявление прямо на семенах их гибридной природы (за счет участия в оплодотворении чужеродной пыльцы) - получило название ксений (от греч. xenia - чужая) (рис. II. 4).
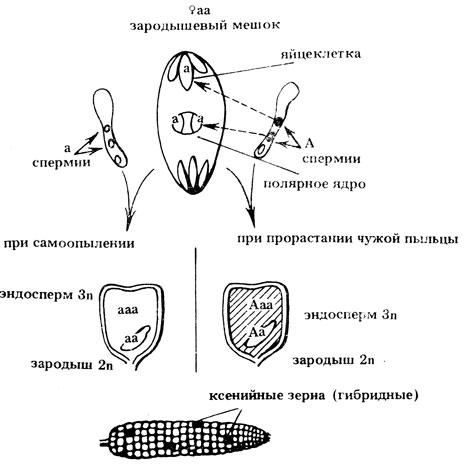
Рис. II.4. Формирование ксений
Если половое размножение у растений осуществляется при инбридинге, то происходит слияние половых клеток родственных растений. Наиболее эффективным способом инбридинга является самоопыление, свойственное большинству видов однолетних растений.
Аутбридинг (неродственное скрещивание) обеспечивается наличием множества приспособлений. Прежде всего это генетические системы, контролирующие несовместимость родственных скрещиваний. Кроме того, разделение полов, разделение во времени созревания мужских и женских гамет, образование на поверхности рыльца защитного слоя, предупреждающего прорастание собственной пыльцы, и другие приспособления обеспечивают регуляцию степени свободного опыления и инбридинга. Растения могут иметь обоеполые цветки, быть однодомными и двудомными. Пестичные и тычиночные цветки иногда собраны в мужские и женские соцветия, как, например, у кукурузы, или распространены по всему растению (огурец) и т. д. У растений с обоеполыми цветками возможно как самоопыление, так и перекрестное опыление; у двудомных - только перекрестное опыление. Для успешного опыления необходимо наличие восприимчивых рылец, причем период восприимчивости может колебаться у растений разных видов от нескольких часов до месяца. Об этом должен знать исследователь, приступающий к постановке опытов по гибридизации растений.
Для гибридизации растений необходимо знать степень самонесовместимости у изучаемого вида и ее причины. Самонесовместимость не связана с нарушением женского и мужского гаметофита. Она проявляется в неспособности прорастания пыльцы на рыльце своего цветка и осуществляется в пестике, который представляет диплоидную спорофитную ткань, контролируемую генотипом спорофита. Системы самонесовместимости (автостерильности) у растений осуществляют генетический контроль оплодотворения и являются приспособлением для перекрестного опыления, обеспечивающего значительную генетическую гетерогенность популяций. Самонесовместимость обнаружена более чем у трех тысяч видов покрытосемянных растений в 80 семействах (среди культивируемых более чем у 40 видов - у ржи, гречихи, клевера, табака, свеклы, капусты и др.). Степень самонесовместимости может быть изменена различными воздействиями и приемами: при старении растений, в результате облучения или воздействия высоким давлением, при опылении цветков в стадии бутонизации у некоторых растений можно увеличить завязываемость семян.
При наличии самонесовместимости проведение генетического анализа обычным гибридологическим методом - получением F2 путем самоопыления гибридов F1 - практически невозможно. Поэтому на перекрестноопыляемых растениях приходится прибегать к применению специальных приемов анализа (Федоров, 1964) или использовать искусственно полученные автофертильные линии (Смирнов, Соснихина, 1984).
Кроме самонесовместимости у растений встречается также перекрестная несовместимость, контролируемая гаметофитными генами. Этот тип несовместимости обеспечивает селективность прорастания определенных типов пыльцы.
Наличие системы несовместимости нарушает случайный характер оплодотворения и может служить причиной отклонений в расщеплениях по генам, сцепленным с генами само- и перекрестной несовместимости (см. гл. III).
Работая с растениями, необходимо знать еще одну их особенность. Поскольку семена формируются из элементов зародышевого мешка материнского растения, первое гибридное поколение по признакам семян (окраска, форма, размеры и т. д.) анализируется на материнском исходном растении, второе - на растениях F1 и т. д. Анализ других признаков растений (наличие или отсутствие опушения, воскового налета, антоциана, формы листовой пластинки и др.) проводится в потомстве растений, выращенных из семян F1, F2 (рис. II.5). Незнание этой особенности может привести к путанице поколений и к неправильным выводам о генотипе исходных растений и характере наследования по разным признакам.
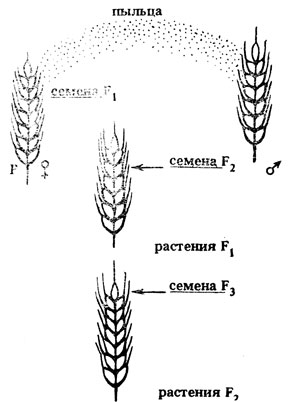
Рис. II.5. Схема анализа признаков семян и растений
У растений при апомиксисе семена образуются либо без слияния гамет из нередуцированной клетки семяпочки, либо из нуцеллуса или зародышевого мешка, возникающего из мегаспоры без мейоза. Очевидно, что в этом случае зародыши будут иметь генотип материнского растения. В результате гаплоидного апомиксиса образуются слабые, обычно стерильные растения, причем потомки гетерозиготного растения могут иметь разные генотипы.
Бесполое (вегетативное) размножение у растений происходит за счет митотических делений клеток. Новый организм развивается из группы специализированных клеток, тканей или органов - боковых побегов, луковиц, корневищ, листьев, усов и т. п. Вегетативное размножение используется в генетическом анализе для размножения мутантов и рекомбинантов, для получения генетически однородных клонов; оно позволяет сохранять в течение длительного времени образцы с идентифицированными генотипами, полиплоидные формы разного типа.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'