Глава III. Наследование при моногенных различиях между исходными формами
III.1. Менделевское наследование. Его модификации
Мендель подчеркивал необходимость тщательного анализа и количественного учета всех потомков в каждом поколении. Поэтому анализ начинается с рассмотрения гибридов первого поколения реципрокных скрещиваний.
Сопоставляя проявление признака в F1 реципрокных скрещиваний, можно определить, связано ли наследование данного признака с хромосомами ядра или с неядерными элементами, сцеплен ли признак с полом; а также выявить гетерозиготность исходных форм, если при постановке скрещиваний генотип родителей не был известен.
Внеядерное наследование проявляется обычно в однородительском типе передачи - гибриды прямого и обратного скрещиваний похожи на мать (матроклинное наследование).
При ядерном наследовании результаты F1 зависят от локализации гена в аутосоме или половой хромосоме; фенотип гибридов может зависеть от условий среды, от пола и т. д.
В случае типичного менделевского наследования, связанного с аутосомной локализацией гена, в реципрокных скрещиваниях гомозиготных форм в F1 проявляется единообразие гибридов. При полном доминировании гибриды F1 похожи на одного из родителей, при неполном наблюдается промежуточное проявление признака, при кодоминировании гибриды могут быть не похожи на обоих родителей. Например, от гомозиготных родителей с группами крови А и В будут рождаться дети с группой крови АВ.
Следует предостеречь начинающего исследователя от ошибки, которая встречается в научной и учебной литературе. Она заключается в том, что на основе фенотипического сходства гибридов F1 с одним из родителей делается вывод о доминировании. Это недоразумение основано, по-видимому, на том, что термин "доминантный" был предложен Менделем для обозначения признака, проявляющегося в гибриде. При этом подразумевалось, что каждый признак находится под контролем одного гена. В дальнейшем был открыт полигенный контроль большинства признаков и термин "доминантный" стали применять только для обозначения аллельных взаимодействий. При полигенном контроле гибриды F1 также могут иметь фенотипическое сходство с одним из родителей, но оно является следствием разных типов неаллельных взаимодействий.
Следовательно, вывод о типе взаимодействия генов - аллельных (разные типы доминирования) или неаллельных (эпистаз, комплементарность, полимерия) может быть сделан только после установления числа генов, по которым идет расщепление, т. е. только после анализа F2.
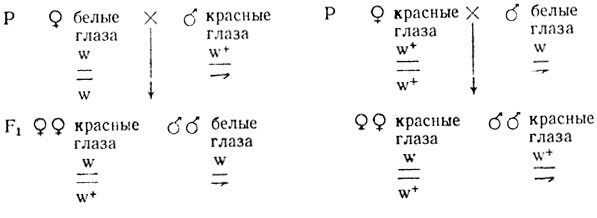
При локализации гена в половых хромосомах - X или Y - результаты F1 реципрокных скрещиваний будут иными. Если признак сцеплен с полом - ген локализован только в Х-хромосоме, то в F1 одного из реципрокных скрещиваний проявляется крисс-кросс наследование - признак от матери переходит к сыну, от отца - к дочери, в другом наблюдается единообразие. Классическим примером такого наследования является наследование белой окраски глаз у дрозофилы, определяемой геном white (w).
Если ген локализован в Y-хромосоме (голандрическое наследование), признак передается особями одного пола и проявляется только у особей того же пола независимо от того, доминантным или рецессивным является этот признак (практически это невозможно установить, так как ген находится в гемизиготном состоянии). Например, признак "темное пятно на спинном плавнике" у гуппи передается по такому типу:
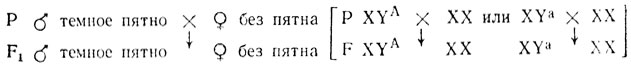
При частичном сцеплении с полом, когда ген локализован в обеих половых хромосомах (X и Y) в F1 реципрокных скрещиваний, наблюдается единообразие, и частичное сцепление с полом обнаруживается только по особенностям расщепления в F2.
Проявление признака у гибрида в некоторых случаях зависит от влияния какого-либо фактора, способного приводить даже к "перемене доминирования". Например, у гороха чувствительность гетерозигот к вирусу желтой мозаики зависит от температуры. Если гетерозиготные растения выращивать при температуре 18° и ниже, то они устойчивы к вирусу, а при 27° и выше - чувствительны к нему. Проявление признака у гомозигот не зависит от температуры (АА всегда чувствительны, аа - устойчивы), и расщепление в F2 будет разным при разной температуре: 3/4 устойчивых: 1/4 чувствительных при 18° и ниже, 3/4 чувствительных: 1/4 устойчивых при 37° и выше.
Проявление ветвистости колоса у гетерозиготных растений некоторых сортов пшениц зависит от продолжительности светового дня - на коротком дне у гетерозигот проявляется ветвистость колоса, на длинном - развивается нормальный колос. Эти примеры показывают, что опыты следует проводить в одинаковых условиях, и объясняют, почему в ряде случаев могут быть получены разные результаты.
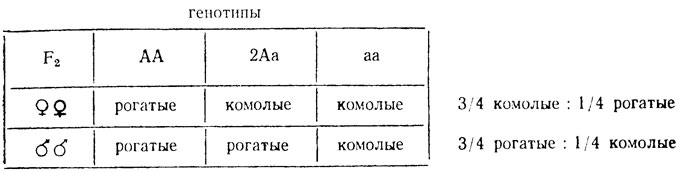
На проявление признаков у гетерозигот могут влиять гормоны. Так зависимые от пола признаки по-разному проявляются у гетерозиготных самцов и самок. Например, у некоторых пород овец рогатость доминирует у самцов, но рецессивна у самок. Вследствие этого гетерозиготные самки оказываются комолыми, а самцы - рогатыми:

Таким образом, изучая проявление признака в первом поколении, можно получить достаточно ценную и необходимую информацию о наследовании, поэтому анализ F1 обязателен (чем часто пренебрегают начинающие исследователи).
Нормальные менделевские расщепления осуществляются при соблюдении ряда условий:
1 - при наличии полового процесса; 2 - гомозиготности исходных форм; 3 - равновероятном образовании и выживании гамет разного генотипа у гетерозигот; 4 - случайном характере оплодотворения или равновероятном участии гамет разного генотипа в оплодотворении; 5-равновероятной выживаемости зигот разного генотипа; 6 - полной пенетрантности гена.
Расщепления также зависят от локализации гена, от типа аллельных взаимодействий, от условий среды.
Характер расщепления, т. е. число и соотношение стенотипических классов в F2, лежит в основе выдвижения гипотез о числе генов, по которым происходит расщепление, типе их взаимодействия и локализации.
При локализации гена в аутосоме нормальное моногенное менделевское расщепление в реципрокных скрещиваниях одинаково - 3/4 доминантных: 1/4 рецессивных потомков при полном доминировании или 1/4:2/4:1/4 при неполном доминировании и кодоминировании.
В случае признаков, сцепленных с полом, результаты реципрокных скрещиваний различны: в одном скрещивании это расщепление 3/4:1/4, причем рецессивный признак проявляется только у особей гетерогаметного пола, а в реципрокном - 1/2:1/2 (независимо от пола):
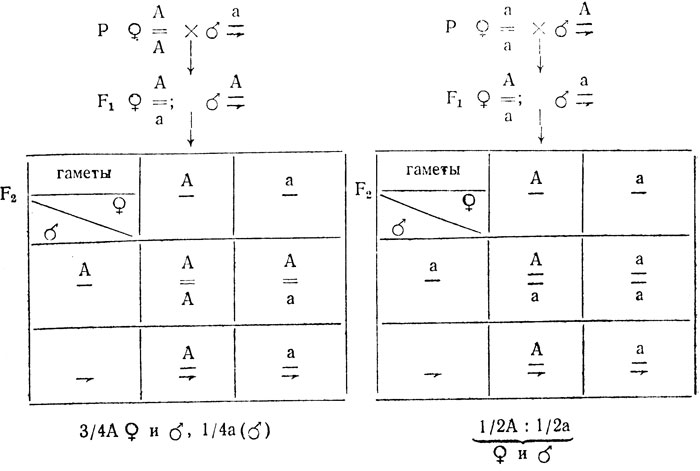
При частичном сцеплении с полом, т. е. при локализации гена как в Х-, так и в Y-хромосоме, расщепления в реципрокных скрещиваниях будут одинаковыми - 3/4А:1/4а, однако рецессивный признак при этом в одном скрещивании проявится у особей женского, в другом - у особей мужского пола:
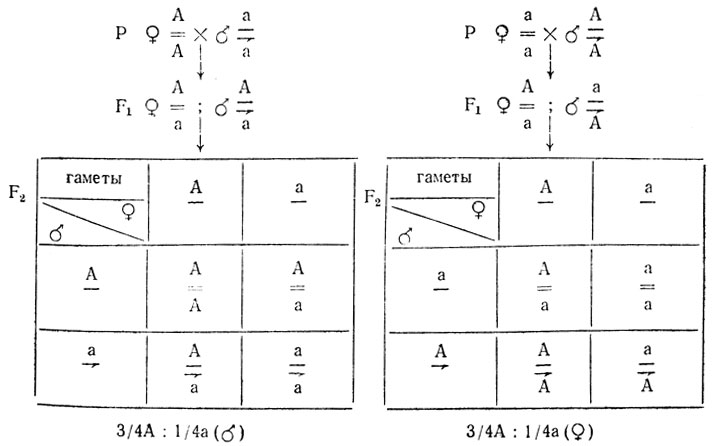
Зависимые от пола признаки так же по-разному проявляются у особей разного пола, хотя характер расщепления в F2 у них будет одинаковым - 3/4:1/4
Мы напомнили эти известные из курса общей генетики типы расщеплений по одному гену, так как они представляют собой возможные "модельные" расщепления. Понятно, что в процессе генетического анализа приходится сталкиваться не только с такими простыми расщеплениями, но и с отклонениями от них. Цель анализа - понять расщепление и на его основе сделать заключение о характере различий между изучаемыми формами, выявить возможные отклонения для последующего исследования их причин.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'