IV.2.1. Расщепления при локализации взаимодействующих генов в аутосоме
Анализ расщеплений при локализации взаимодействующих генов в одной аутосоме усложняется в силу ряда причин. Прежде всего по расщеплению в F2 не всегда можно определить силу сцепления между генами; в ряде случаев устанавливают только факт сцепления. Это связано с тем, что в F2часть потомков, возникающих с участием кроссоверных гамет, фенотипически не отличается от некроссоверных. Вероятность выщепления двойных рецессивов зависит при этом от типа гетерозиготы. В потомстве цис-гетерозигот (гены в "притяжении") двойные рецессивы возникают от слияния некроссоверных гамет и их частота тем больше, чем сильнее сцеплены гены. В потомстве транс-гетерозигот (гены в "отталкивании") двойные рецессивы выщепляются значительно реже, только при слиянии двух кроссоверных гамет. Их частота при сильном сцеплении может быть очень мала, или они вообще не выщепляются в небольших выборках.
Это можно продемонстрировать с помощью решеток Пеннета, в которых отражены доли кроссоверных и некроссоверных гамет для случая, когда расстояние между генами равно 10% кроссинговера и кроссинговер идет у самцов и самок с одинаковой частотой.
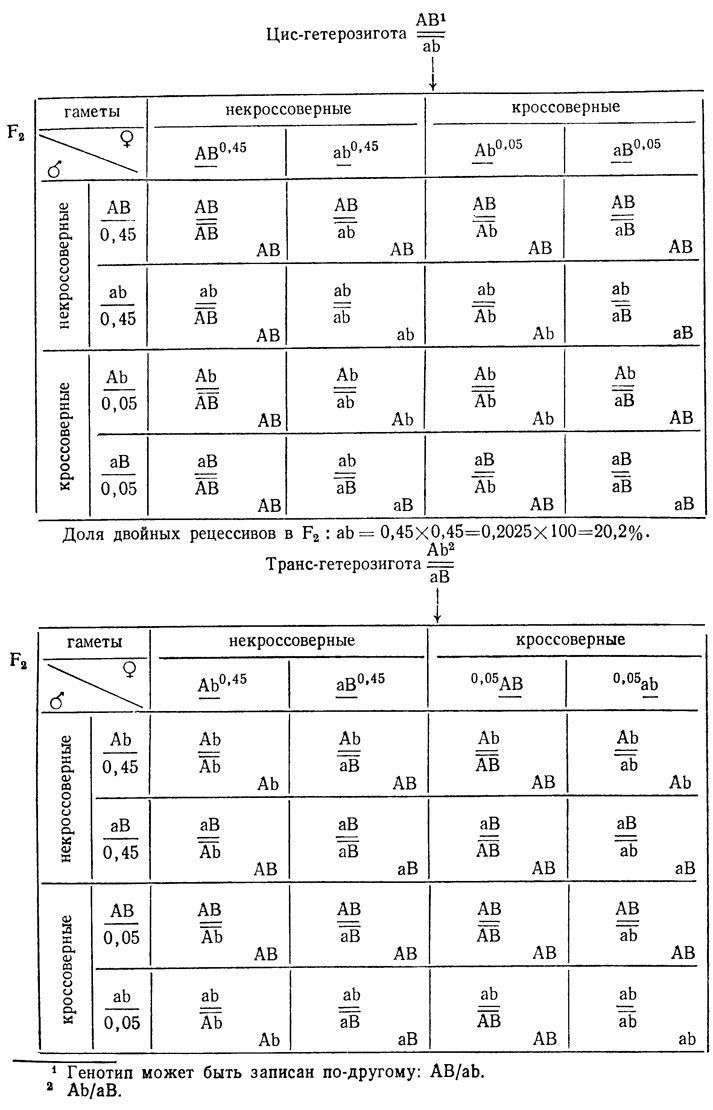
Доля ab = 0,05×0,05 = 0,0025×100 = 0,25%. В обоих случаях часть классов, возникших с участием кроссоверных гамет, по фенотипу неотличима от некроссоверных.
Сложность анализа связана также с невозможностью выделения рекомбинантных классов, если их фенотип при взаимодействии между генами не отличается от фенотипа некроссоверов. Наконец, неодинаковая частота кроосинговера у особей разных полов затрудняет анализ сцепленного наследования взаимодействующих генов и определение процента кроссинговера между ними.
В некоторых случаях удается преодолеть эти трудности, применив логический анализ.
Для проверки аллельности двух независимо возникших рецессивных мутаций, вызывающих светло-зеленую окраску растений гороха, мутантов скрестили между собой. В F1 все растения оказались зелеными, а в F2 произошло расщепление: 289 зеленых и 131 светло-зеленых. Аллельны ли мутации? Объяснить расщепление и определить генотипы исходных форм.
Поскольку гетерозиготные растения F1, полученные от скрещивания двух рецессивных мутантов, оказались зелеными (дикий тип), можно утверждать, что мутации неаллельны. Следовательно, изучаемые мутанты различаются, по крайней мере, по двум генам. Расщепление на два класса свидетельствует об их взаимодействии. Если бы эти гены наследовались независимо, то можно было ожидать расщепления 9:7, поскольку в F1 наблюдается эффект комплементации. Однако проверка по методу χ2 отвергает гипотезу о независимом наследовании генов - χ2 = 27,0, p<<0,01. Очевидно, гены сцеплены. Зеленая окраска проявляется только при наличии в зиготе доминантных аллелей обоих генов - АВ, во всех остальных случаях - Аb, аВ и ab - окраска должна быть светло-зеленая. Гены вводились в скрещивание в транс-положении. При их сильном сцеплении в отсутствии кроссинговера расщепление должно было быть 1 зеленый:1 светло-зеленый:
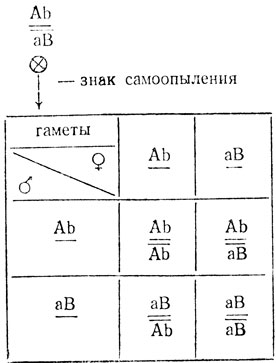
Поскольку расщепление в опыте не 1:1, можно предполагать, что в гетерозиготе имел место кроссинговер. Однако в этом случае некроссоверные и кроссоверные классы неразличимы фенотипически.
Если бы в коллекции имелся анализатор по этим генам - двойной рецессив aabb, то по результатам анализирующего скрещивания можно было бы определить процент кроссинговера, так как зеленые растения в этом случае могли бы появиться только в результате кроссинговера:
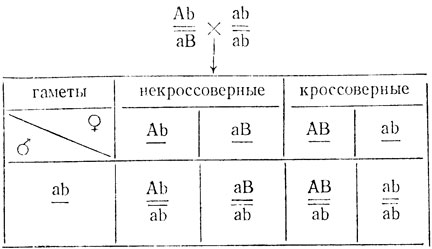
Особи с фенотипом АВ - зеленые, остальные (Аb, аВ и ab) - светло-зеленые. Поскольку частоты рекомбинантов АВ и ab должны быть примерно одинаковы, процент кроссинговера можно вычислить по формуле 2×m/n×100%, где m - число кроссоверных зеленых растений АВ; n - общее число растений в потомстве.
Используя в анализирующем скрещивании гибридную форму в качестве материнского или в качестве отцовского растений, можно установить, одинаковы ли частоты кроссинговера у особей разного пола. Взаимодействие сцепленных генов приводит к уменьшению числа классов в расщеплении, и частоту кроссинговера можно косвенно определить по доле проявившихся кроссоверов (или по доле некроссоверных классов).
Если кроссинговер идет только у особей одного пола, то сцепление по результатам F2 обнаруживается при скрещивании любых гетерозигот. Однако определение процента кроссинговера возможно только при скрещивании цис-гетерозигот. Он равен удвоенной доле фенотипически проявившихся кроссоверов:
так как половина особей, возникших с участием кроссоверных гамет, будет фенотипически неотличима от некроссоверных:
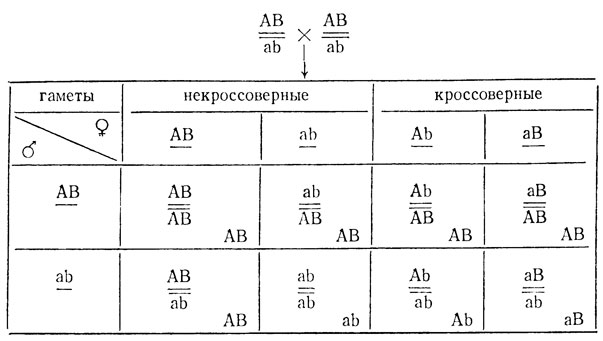
В потомстве транс-гетерозигот кроссоверы вообще не проявляются, при этом соотношение фенотипов в расщеплении 1аВ:2АВ:1Аb, если АЬ и аВ фенотипически различимы, или 1АВ:1(Аb + аВ), если Аb и аВ неразличимы. Наконец, оно может быть 3(АВ + Аb) (или аВ):1аВ (или Аb), если АВ не отличается от Аb или от аВ
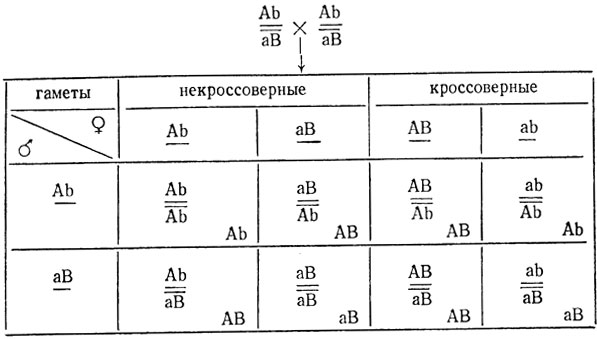
В реципрокных скрещиваниях дрозофил с красными и белыми глазами были получены одинаковые результаты: все мухи F1 оказались красноглазыми, а в F2 произошло расщепление: 273 с красными, 54 с белыми, 53 с коричневыми и 41 с ярко-красными глазами (2=421). При скрещивании гомозиготных мух с ярко-красными и коричневыми глазами все потомки F1 имели красные глаза, а в F2 произошло расщепление: 118 с красными, 50 с ярко-красными и 48 с коричневыми глазами. Объяснить расщепления и определить генотипы исходных мух и гибридов F1 в обоих скрещиваниях.
Из F1 реципрокных скрещиваний можно сделать заключение, что признак не сцеплен с полом и исходные особи, по-видимому, гомозиготны. Расщепление на четыре фенотипических класса в F2 свидетельствует об участии в расщеплении не менее двух взаимодействующих генов. Это расщепление явно отличается от теоретически ожидаемого при независимом наследовании 9:3:3:1 - (236,9:78,9:78,9:26,3), причем в расщеплении преобладают классы, сходные с родительскими формами, что позволяет предположить сцепленное наследование двух генов. Н0 - произошло расщепление по двум сцепленным взаимодействующим генам, локализованным в аутосоме. Гибриды F1 так же, как и один из родителей, красноглазы. По-видимому, генотипы красноглазых гибридных особей F1 - AB/ab. Тогда появление четырех фенотипических классов в F2 можно объяснить кроссинговером, в результате которого появились новые (по сравнению с родительскими) классы мух - ярко-красноглазые и коричневоглазые. Поскольку у самцов дрозофилы кроссинговер не происходит, то в F2 проявляется только половина кроссоверов. Следовательно, примерный процент кроссинговера между исследуемыми генами равен:
Результаты скрещивания мутантных мух с коричневыми и ярко-красными глазами подтверждают высказанное предположение: потомки F1 имеют красные глаза (дикий тип). Следовательно, мутации неаллельны и в F1 образовались транс-гетерозиготы - Аb/аВ.
Из-за отсутствия кроссинговера у самцов кроссоверные классы в этом скрещивании в F2 не проявляются, и расщепление должно идти по формуле 1Аb:2АВ:1аВ, χ2 = 1,88, p>0,50. Отклонение в опыте случайно, гипотеза не отвергается.
Исходные формы различаются по двум комплементарно взаимодействующим генам, локализованным в аутосоме. Красная окраска глаз проявляется только при наличии в зиготе доминантных аллелей обоих генов - АВ, белая - рецессивных аллелей этих генов (ab), коричневая - одним из генов в рецессиве (Аb или аВ), ярко-красная соответственно аВ или Аb. Расстояние между генами 44,6±2,4% кроссинговера. Генотипы исходных мух: красноглазые - АВ/АВ, белоглазые - ab/ab, генотип гетерозигот F1 - AB/ab. Ярко-красноглазые Ab/Ab, коричневоглазые аВ/аВ, генотип гетерозигот F1 - Аb/аВ.
Для проверки правильности сделанных выводов следует провести дальнейший генетический анализ. Например, можно поставить три анализирующих скрещивания: 1 - ♀AB/ab×♂ab/ab, 2 - ♂Ab/aB×♀ab/ab, 3 - ♀AB/ab×♂ab/ab. В первом скрещивании должны появиться красноглазые и белоглазые мухи в соотношении - 1:1; во втором - также два класса мух с ярко-красными и коричневыми глазами (~1:1). В обоих скрещиваниях у гетерозиготных самцов не идет кроссинговер. В третьем скрещивании в случае кроссинговера между генами появятся четыре фенотипических класса, и если процент кроссинговера по результатам F2 был определен достаточно точно, то можно ожидать следующего соотношения фенотипов в расщеплении.
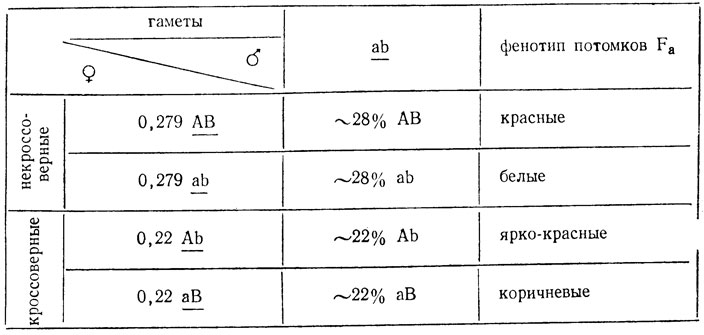
По соотношению фенотипов в третьем скрещивании можно уточнить процент кроссинговера.
В реципрокных скрещиваниях дрозофил с красными и темно-коричневыми глазами в F1 были получены красноглазые мухи, а в F2 произошло расщепление: 285 с красными, 97 с темно-коричневыми и 16 с ярко-красными глазами (∑ = 398). В возвратном скрещивании гибридных самок F1 с коричневоглазыми исходными самцами получили расщепление: 71 с красными, 90 с темно-коричневыми и 15 с ярко-красными глазами (∑ = 176). Объяснить расщепления, определить генотипы исходных мух и гибридов F1.
Результаты F1 реципрокных скрещиваний говорят о том, что признак не сцеплен с полом и исходные мухи гомозиготны. Расщепление в F2 не соответствует формуле моногенного расщепления 1:2:1. (χ2 = 107,2, p<<0,01) Н0 - в расщеплении участвуют два взаимодействующих гена. Оно отличается от формул независимого наследования двух генов - не 9:3:4, (χ2 = 26,2, p<<0,01); не 12:3:1, (χ2 = 10,2, p<<0,01); не 9:6:1, (χ2 = 38,2, p<<0,01), причем среди потомков F2 преобладают особи с фенотипами исходных мух. Очевидно, гены сцеплены. Гибриды F1, так же как один из родителей, имеют красные глаза (дикий тип). Тогда генотипы исходных красноглазых мух АВ/АВ, мух с темно-коричневыми глазами ab/ab, потомков F1-AB/ab. Появление в F2 трех фено-типических классов свидетельствует о кроссинговере, прошедшем у гетерозиготных самок F1, а также о том, что особи одного из кроссоверных классов (например, АЬ) фенотипически не отличаются от двойного рецессива, а второго имеют ярко-красные глаза (аВ):
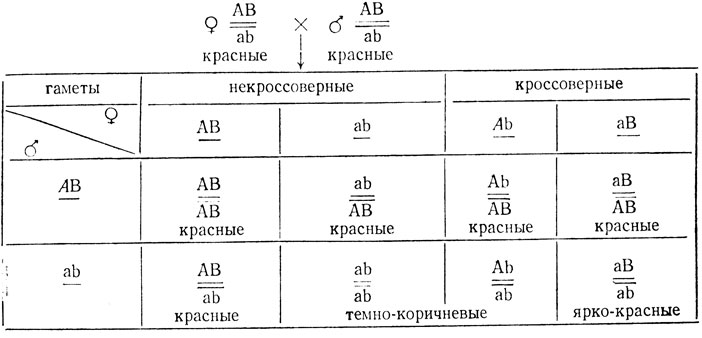
Из решетки видно, что мухи с ярко-красными глазами (аВ) составляют 1/4 часть всех потомков, возникших с участием кроссоверных гамет. Следовательно, можно грубо оценить процент кроссинговера, помножив долю ярко-красноглазых мух (16/398 = 0,04) на четыре: 0,04×4×100% = 16% ± 1,8%.
Возвратное скрещивание в данном случае является анализирующим, наличие трех фенотипических классов в его потомстве подтверждает участие в расщеплении двух генов, а соотношение классов свидетельствует о сцеплении генов и прошедшем кроссинговере. Кроссоверные ярко-красноглазые мухи составляют в Fa1/2 от всех кроссоверов.
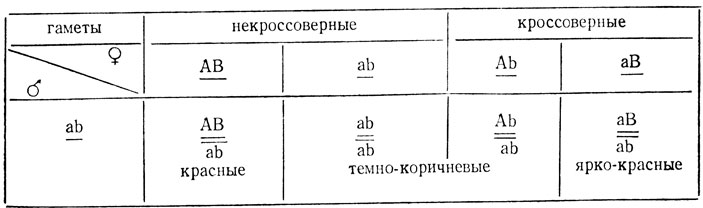
Отсюда следует, что процент кроссинговера равен 15/176×2×100% = 17% ±2,8%, что хорошо согласуется с результатами, полученными в F2.
Исходные формы различаются по двум взаимодействующим сцепленным генам, локализованным в аутосоме на расстоянии 16,5±2,3% кроссинговера (16+ 17/2×100 = 16,5%). Гены взаимодействуют по типу комплементарности: АВ - красная окраска глаз, Аb и ab - темно-коричневая, аВ - ярко-красная. Генотипы исходных мух: красноглазых АВ/АВ, темно-коричневоглазых ab/ab, гибридов F1-AB/ab.
Окончательным доказательством сцепления генов могло бы служить скрещивание гетерозиготных самцов из F1 с самками из исходной линии мух с темно-коричневыми глазами. В таком скрещивании - ♂AB/ab×♀ab/ab должно было бы появиться два класса мух в равном количестве - 1/2 красноглазых и 1/2 с темно-коричневыми глазами, так как у самцов не идет кроссинговер.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'