V.2.2. Случайное хроматидное расщепление. Двойная редукция
Случайное хроматидное расщепление у автополиплоидов осуществляется в том случае, когда участок от гена до центромеры достаточно велик для прохождения на нем кроссинговера (центромера и ген не проявляют сцепления). Если происходит кроссинговер, то хроматиды, исходно удерживаемые вместе одной центромерой, могут оказаться соединенными с разными центромерами (рис. V.4). Очевидно, что хроматиды с аллелями а22 и а31, прикрепленные до кроссинговера ко второй и третьей центромерам соответственно, после прохождения кроссинговера соединяются с другими центромерами: а22 с третьей (III), а31 со второй (II). Вследствие этого при попарном расхождении хромосом аллели а21 и а22 могут оказаться в одной гамете, то же - аллели а31 и а32. К таким же результатам приведет кроссинговер с участием разных хроматид 1-3, 1-4 и 2-4 хромосом. Следовательно, при попарном расхождении хромосом все восемь хроматид в анафазе второго деления мейоза могут распределяться случайно, и все возможные пары из восьми хроматид имеют равную вероятность попасть после двух мейотических делений в одну гамету. Число их возможных сочетаний (С) при этом равно 28, что легко определить по формуле: Cmn = n!/(n-m)!m!, где n = 8 число хроматид в квадриваленте, m = 2, так как расхождение попарное. Соотношение гамет разного типа зависит от генотипа тетраплоида. У триплекса АААа частота гамет АА-С62 = 15, аа-С22 = 1, Аа-28-16 = 12; у дуплекса ААаа частота гамет АА-С42 = 6, аа-С42 = 6, Аа-28-12 = 16 и т. д.
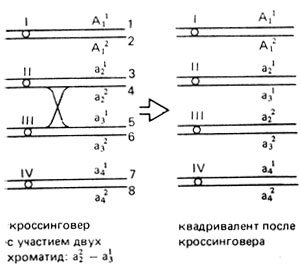
Рис. V.4. Схема кроссинговера в квадриваленте с участием двух хроматид
В результате случайного хроматидного расщепления тетраплоиды с генотипом триплекса АААа и симплекса Аааа могут образовывать некоторое количество гамет аа и АА соответственно. Это происходит в случае нерасхождения аллелей, локализованных в сестринских хроматидах, если между геном и центромерой произойдет кроссинговер. Такое явление получило название двойной редукции. Частота двойной редукции зависит от расстояния между геном и центромерой. Наибольший процент двойной редукции наблюдается в случае, когда это расстояние равно 50% кроссинговера и в нем участвуют четыре хроматиды (рис. V.5). Аналогично может идти кроссинговер с участием других хроматид.
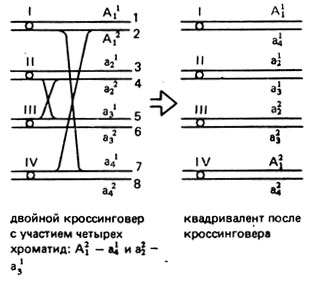
Рис. V.5. Схема кроссинговера в квадриваленте с участием четырех хроматид
Очевидно, что при 50% кроссинговере между геном и центромерой нарушается случайное комбинирование хроматид в гаметах, так как 4 комбинации хроматид содержат по две сестринских хроматиды, т. е. при двойной редукции образуется не 28, а 24 типа гамет, и их соотношение несколько отличается от соотношения гамет при случайном хроматидном расщеплении (табл. V.2).

Таблица V.2. Возможные варианты попарного расхождения хромосом в первом и хроматид во втором делении мейоза при условии 50% кроссинговера на участке между геном и центромерой с участием 4 хроматид в симплексе
Примечание: Подчеркнуты гаметы, возникшие в результате двойной редукции. Соотношение гамет - 1АА:10Aa:13aa.
Теоретически рассчитано, что при кроссинговере между геном и центромерой, равном 42,85%, соотношение гамет и фенотипов соответствует случайному хроматидному расщеплению.
В табл. V.3 представлены соотношения гамет и расщепления в потомстве от скрещивания автополиплоидов с разными генотипами при случайном хроматидном расщеплении и при двойной редукции.
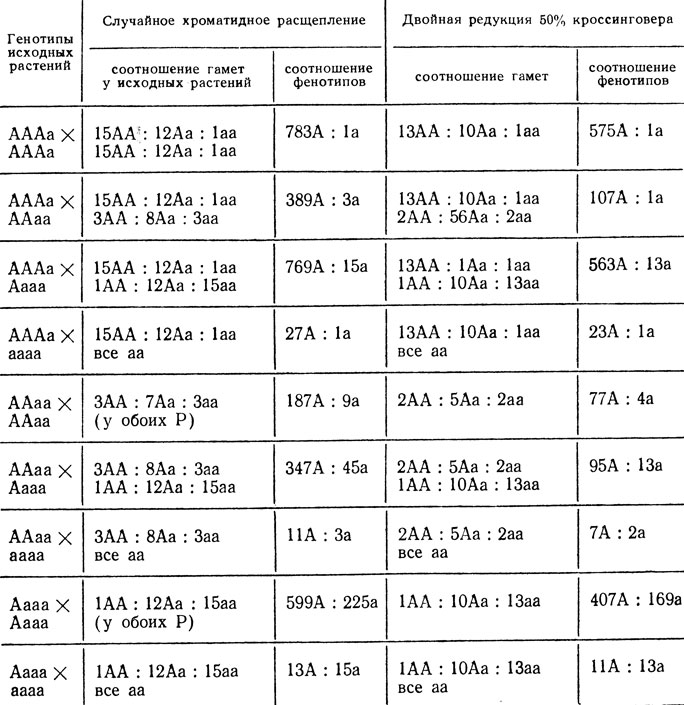
Таблица V.3. Соотношение гамет и фенотипов, возникающих при хроматидном расщеплении аутотетраплоидов при полном доминировании
Таким образом, если ген локализован вблизи центромеры, то осуществляется, как правило, случайное хромосомное расщепление, если далеко от центромеры - в основном случайное хроматидное. Это усложняет генетический анализ, когда изучают наследование двух генов, локализованных на разных расстояниях от центромеры. Следует отметить большое разнообразие потомков по генотипу и однородность, сходство их по фенотипу, а также малую вероятность выщепления рецессивных гомозигот.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'