VII.3. Определение группы сцепления у грибов-аскомицетов на основе гаплоидизации
У многих видов мицелиальных грибов обнаружен парасексуальный цикл, приводящий к появлению генетических рекомбинантов без участия мейоза. В процессе этого цикла образуются гетерокарионы, содержащие в мицелии ядра двух разных штаммов, например ауксотрофных по разным факторам роста (см. гл. II). Они легко выделяются на минимальной среде, так как растут на ней, в то время как родительские штаммы не растут. При слиянии в гетерокарионе двух ядер возникает диплоидное ядро, образующее при размножении (митозе) диплоидные клетки. В потомстве гетерозиготных диплоидов могут появляться рекомбинанты (сегреганты) по рецессивным маркерам. Их выщепление может быть следствием митотического кроссинговера (см. с. 215) или результатом нерасхождения хромосом. Например, при гаплоидизации в диплоидной культуре аспергилла, гетерозиготного по гену окраски конидий (ω/ω+), может появиться белый сектор. Это результат потери хромосомы, в которой локализован аллель ω+, обусловливающий зеленую окраску конидий. На мицелии в белом секторе образуются гаплоидные конидии, которые отличаются по размеру от диплоидных (по диаметру они в 1-1,4 раза меньше), что хорошо видно под микроскопом (рис. VII.4).
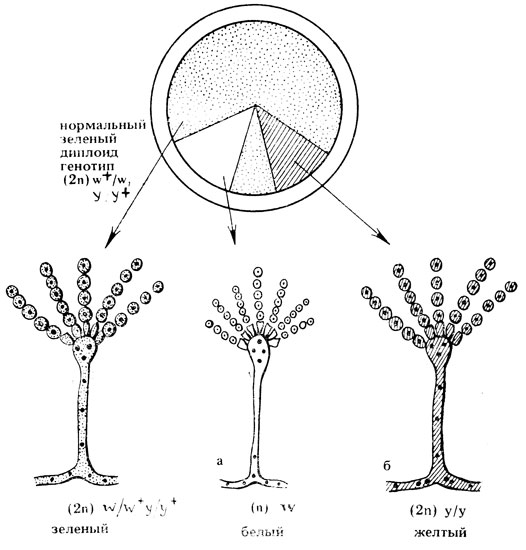
Рис. VII.4. Выщепление (сегрегация) рецессивных мутаций и аспергилла впоследствие гаплоидизации и митотического кроссинговера. Генотип диплоидной культуры аспергилла +/w, +/y (w - белые колонии, y - желтые колонии), а - выщепление белых гаплоидных конидий вследствие гаплоидизации (аналогично может происходить сегрегация желтых гаплоидных конидий), б - выщепление желтых диплоидных конидий вследствие кроссинговера при парасексуальном процессе (такая же сегрегация может приводить к образованию белых диплоидных конидий)
Нерасхождение приводит к утрате одной из хромосом и к образованию ядер с несбалансированным набором хромосом - трисомиков (2n+1) и моносомиков (2n-1). Это состояние неустойчиво и может приводить к новым нерасхождениям, которые у моносомиков ведут к потерям других хромосом в последующих митозах и в конечном итоге к утрате одного из гомологов по всем парам хромосом, т. е. к гаплоидизации. При этом не происходит рекомбинации между генами, локализованными в одной хромосоме, т. е. эти гены обнаруживают полное сцепление. Гены разных хромосом рекомбинируют с частотой 50%. Например, если гены АВ находятся в одной, а гены CDE - в другой хромосоме, то у гаплоидов, возникших в результате гаплоидизации от диплоидного гибрида с генотипом 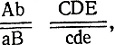 могут быть генотипы:
могут быть генотипы: 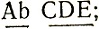
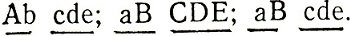 Отсутствие рекомбинаций между генами АВ, а также между генами CDE, свидетельствует о сцеплении генов АВ и CDE. Наличие 50% рекомбинантов между этими группами указывает на их принадлежность к разным хромосомам.
Отсутствие рекомбинаций между генами АВ, а также между генами CDE, свидетельствует о сцеплении генов АВ и CDE. Наличие 50% рекомбинантов между этими группами указывает на их принадлежность к разным хромосомам.
У плесневых грибов спонтанная гаплоидизация происходит редко, но ее частота может быть увеличена при добавлении в среду некоторых веществ, например, n-фторфенилаланина, которым обрабатывают диплоидные клоны. У дрожжей-сахаромицетов нет полной гаплоидизации, но могут происходить потери отдельных хромосом. Гаплоидизация и потери хромосом используются в генетическом анализе при определении группы сцепления у мицелиальных грибов.
Определение группы сцепления - одна из задач локализации гена. Вторая задача - установление места нахождения гена в группе сцепления и картирование хромосом (см. гл. VIII).
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'