VIII.4. Картирование на основе полиморфизма (длин рестрикционных фрагментов ДНК (ПДРФ). Физические карты хромосом
Открытие специфических рестрикционных нуклеаз (рестриктаз), узнающих специфические последовательности нуклеотидов и разрезающих ДНК на фрагменты, стало одной из предпосылок картирования генома на основе ПРДФ. Разные рестриктазы разрезают ДНК в разных сайтах, для каждой из них количество и локализация сайтов рестрикции строго определены. С помощью рестриктаз можно разрезать ДНК на фрагменты определенной длины с точностью до одного нуклеотида.
Например, ДНК бактериальных плазмид может быть разрезана всего на несколько фрагментов (рестриктов). Размер фрагментов однозначно устанавливается с помощью электрофореза в агарозном геле: чем меньше фрагмент, тем больше его электрофоретическая подвижность в геле. На одну из дорожек геля наносят стандартные фрагменты ДНК известного размера и с их помощью определяют длину исследуемых фрагментов ДНК. Это обычный метод, применяемый для разделения, идентификации и очистки фрагментов ДНК (рис. VIII. 12). На основании учета длины рестрикционных фрагментов строят рестрикционные карты ДНК данной плазмиды. Рестрикционной или физической картой называют схему, на которую нанесены сайты узнавания разных рестриктаз, дана линейная последовательность этих сайтов и расстояние между ними в парах нуклеотидов (п. н. для коротких отрезков, т. п. н.- тысячи - для длинных отрезков).
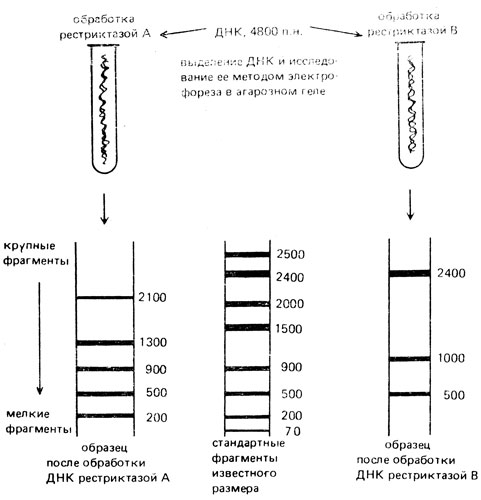
Рис. VIII.12. Определение длины стандартных фрагментов ДНК, возникающих под действием разных рестриктаз
Для составления рестрикционной карты используют методы: последовательного расщепления ДНК двумя рестриктазами, частичное расщепление ДНК, меченой радиоактивной меткой по определенному концу и др. Для примера рассмотрим способ построения рестрикционной карты путем последовательного расщепления ДНК двумя рестриктазами. При этом способе сначала определяют величины фрагментов, образуемых под действием одной рестриктазы (I). Затем полученные фрагменты обрабатывают другой рестриктазой (II). И наоборот, фрагменты, полученные под действием рестриктазы II, обрабатывают рестриктазой I. На основе логического анализа, сопоставляя размеры фрагментов, образовавшихся после этих расщеплений, строят рестрикционные карты линейной или кольцевой ДНК.
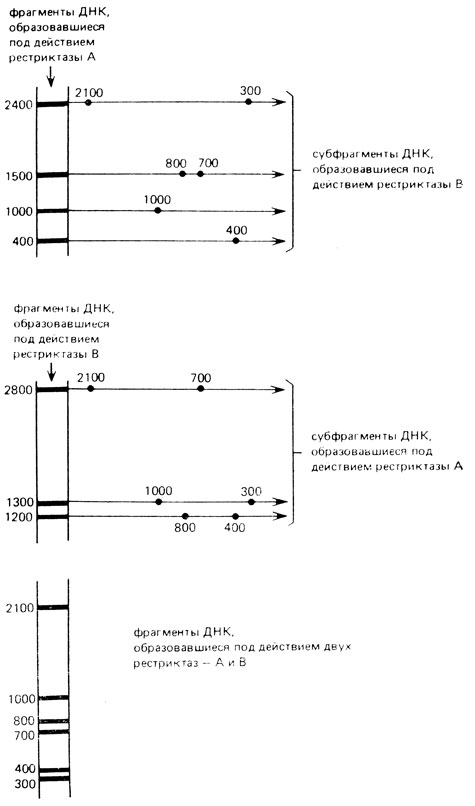
Рис. VIII.13. Расщепление ДНК на фрагменты двумя рестриктазами. Анализ с помощью двухмерного электрофореза
Рассмотрим построение рестрикционной карты участка линейной ДНК 5500 п. н. На рис. VIII. 13 представлены размеры фрагментов, полученных после последовательной обработки ДНК двумя рестриктазами, А- и В-методом двухмерного электрофореза. Из рисунка видно, что наибольший фрагмент, полученный после обработки ДНК рестриктазой А (А-2400), разрезается рестриктазой В на два фрагмента - 2100 п. н. и 300 п. н. Наибольший фрагмент, полученный под действием рестриктазы В (В - 2800 п.н.), разрезается рестриктазой А тоже на два фрагмента - 2100 и 700 п. н. Каждый фрагмент, полученный из исходных А-фрагментов под действием фермента В, есть в образцах, содержащих фрагменты, полученные в результате расщепления исходных В-фрагментов рестриктазой А. В геле, содержащем продукты двойного расщепления, каждый фрагмент встречается один раз. Используя принцип "перекрывающихся фрагментов" можно построить карту этого района.
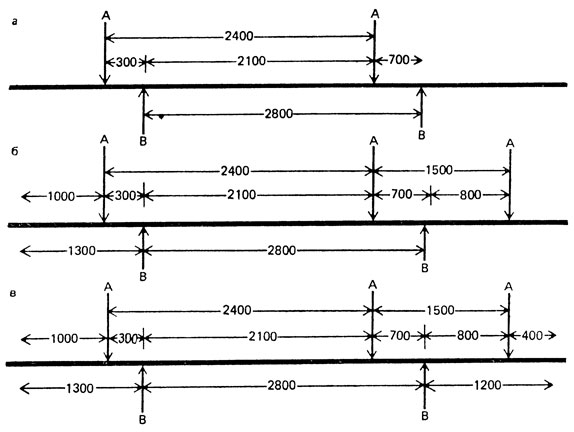
Рис. VIII. 14. Последовательность построения рестрикционной карты фрагмента линейной ДНК 5500 п. н. (а, б, в)
В нашем примере фрагмент 2100 п.н. образуется в результате расщепления фрагментов А-2400 и В-2800, т. е. они перекрываются и область 2100 п. н. для них общая. С одной стороны этого фрагмента на расстоянии 300 п.н. находится А-сайт рестрикции,с другой - на расстоянии 700 п. н. - В-сайт. На основе этих данных можно нарисовать участок карты, обозначив на ней сайты рестрикции рестриктаз А и В вертикальными черточками и размеры фрагментов между ними (рис. VIII. 14, а). Затем можно продолжить карту вправо и влево, выяснив, из каких фрагментов образовались участки 700 и 300 п. н. Фрагмент 700 п. н. образуется при расщеплении фрагмента А-1500 рестриктазой В, следовательно, справа на карте лежит А-сайт на расстоянии 800 п. н. Фрагмент 300 п.н. происходит или из фрагмента В-1300, или из фрагмента А-1000. Поскольку фрагмент А-1000 не разрезается рестриктазой В, он должен находиться с левого края карты (рис. VIII. 14, б). Наконец, фрагмент 800 п.н. получен при расщеплении фрагмента В-1200 под действием рестриктазы А, следовательно, он расположен справа. Поскольку рестриктаза В не расщепляет оставшийся фрагмент 400 п. н., он является правым концом карты данного участка (рис. VIII. 14, в). Карта, построенная на основе этих данных представлена на рис. VIII. 15. Для рестрикциионных карт характерна аддитивность. При рестрикционном картировании можно обнаруживать делеции и дупликации - они приводят к уменьшению или увеличению какого-либо фрагмента по сравнению с нормой (рис. VIII. 16). Для фаговых, хлоропластных и митохондриальных ДНК, размер генома которых относительно невелик и варьирует от нескольких до сотен кб, построение ре-стрикционных карт - обычная процедура.
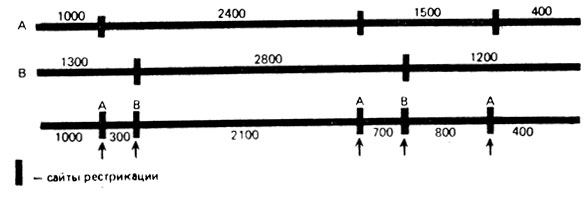
Рис. VIII. 15. Рестрикционная (физическая) карта фрагмента линейной ДНК 5500 п. н
Однако геномы эукариот слишком сложны для такого картирования, так как под действием рестриктаз образуется огромное количество (млн) фрагментов. Это обстоятельство исключает возможность построения рестрикционных карт эукариот на основе анализа рестрикционных фрагментов тотальных препаратов ДНК. Уже на первых этапах изучения структуры клонированных генов эукариот был обнаружен высокий межиндивидуальный полиморфизм последовательностей нуклеотидов ДНК. Он проявляется в том, что рестрикционные карты соответствующих аллелей могут различаться по сайтам рестрикции, т. е. это полиморфизм длины рестрикционных фрагментов (ПДРФ). Например, он был обнаружен при рестрикционном картировании глобиновых генов человека, локализованных в коротком плече хромосомы 11. При обработке этой области генома разными рестриктазами аллельные варианты гена, контролирующего р-полипептид, обнаруживаются в разных фрагментах. Аналогичное явление показано и на аллелях других генов. ПДРФ проявляется даже в том случае, когда изменение нуклеотидной последовательности не обнаруживается фенотипически: он проявляет менделевское моногенное наследование по типу кодоминирования. ПДРФ может быть связан с точковыми мутациями, затрагивающими сайты узнавания для разных рестриктаз, с микроперестройками - делециями или вставками мобильных уникальных последовательностей ДНК, а также с транспозициями мобильных генетических элементов. Очевидно, что уникальные последовательности ДНК (уМГМ) можно использовать как инструмент для выявления ПДРФ. Для этого рестрикционные фрагменты изучают в блот-гибридизации с радиоактивным зондом какого-либо клонированного уникального гена (или уникального фрагмента, функция которого неизвестна). На радиоавтографе у разных индивидуумов могут выявляться фрагменты ДНК разных размеров, характеризующие их межаллельные различия. Если области, гомологичные изучаемому уникальному фрагменту, идентичны по распределению сайтов узнавания для исследуемой рестриктазы, то на радиоавтографе число и положение полос гибридизации будет совпадать.
На основе наличия полиморфных вариантов можно проводить генетический анализ, в том числе и генетическое картирование, используя классический гибридологический метод, с той разницей, что для определения этих различий изучают структуру фрагментов ДНК, а не признаки, альтернативно проявляющиеся фенотипически.
Если наследование какого-либо признака, установленное на основе изучения родословных, совпадает с наследованием МГМ, выявленным на основе рестрикционного картирования соответствующей области генома, то это свидетельствует о сцеплении изучаемого гена с данным МГМ. Например, сравнивая рестрикционные карты гена больного и здорового человека, можно обнаружить наличие определенного сайта рестрикции у больных, что будет свидетельствовать о сцеплении гена с идентифицируемым МГМ. Так у человека был картирован ген, мутация в котором вызывает тяжелое наследственное заболевание - хорею Хетингтона.
Для анализа участков эукариотических хромосом длиной в несколько т. п. н. необходимо клонирование последовательности ДНК. Но фрагменты такой длины невозможно клонировать с помощью фаговых или космидных векторов.
Задача построения рестрикционных карт отдельных участков генома эукариот может быть решена с помощью двух методов.
Первый - это "прогулка по хромосоме". Для картирования этим методом используют несколько рекомбинантных фагов (или космид), содержащих перекрывающиеся фрагменты одного участка ДНК. Сначала выделяют клон, содержащий фрагмент с интересующей нас областью (или вблизи нее), а затем с помощью гибридизации с зондом из библиотеки генома выделяют другие клоны, содержащие последовательности, перекрывающиеся с последовательностью первого клона. Эти клоны содержат, кроме того, последовательности, расположенные справа или слева от фрагмента, содержащегося в первом клоне.
Для выделения каждого последующего клона в качестве зонда используют концевой субфрагмент ДНК, не перекрывающий уже отклонированные последовательности. Затем проводят рестрикционное картирование фрагментов каждых двух клонов и определяют, в каком направлении увеличен фрагмент одного клона по отношению к другому. Этот процесс может быть повторен многократно, в результате чего можно проанализировать последовательности сотен тысяч пар оснований и построить рестрикционную карту данного участка (рис. VIII. 17).
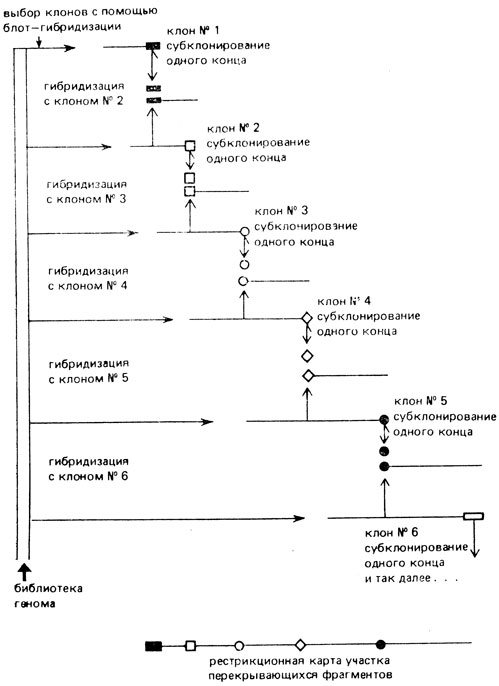
Рис VIII.17. Рестрикционное картирование фрагмента ДНК методом 'прогулки по хромосоме'
Второй метод основан на использовании редкощепящих рестриктаз и пульс-электрофореза, с помощью которых удается выявлять и анализировать фрагменты ДНК длиной до нескольких тысяч кб. С помощью блот-гибридизации можно локализовать на этих фрагментах ДНК любые клонированные последовательности, уже определенные с помощью цитологического или генетического картирования. Затем на основе рестрикционного картирования устанавливают относительное расположение разных клонированных последовательностей (генов) внутри исследуемого фрагмента. Точность такой карты ограничена разрешающей способностью генетического анализа.
В результате применения комплекса всех этих методов строят рестрикционную (физическую) карту хромосомы, в которой определено местоположение генетических, цитологических и молекулярно-генетических маркеров. Определение полной нуклеотидной последовательности этой части генома (секвенирование) - завершающий этап картирования.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'