§ 2. Химическое строение и свойства нуклеиновых кислот
Нуклеиновые кислоты обладают способностью растворяться в воде и щелочах, они могут быть осаждены из раствора кислотами. Биологические полимеры - нуклеиновые кислоты, распадаясь, образуют мономеры - нуклеотиды. Структурными единицами каждого нуклеотида являются фосфат (остаток фосфорной кислоты), углеводный остаток сахара, содержащего пять атомов углевода,- дезоксирибозы (в ДНК) или рибозы (в РНК).
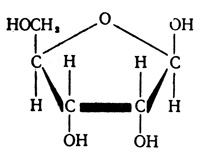
Рибоза
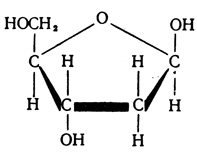
Дизоксирибоза
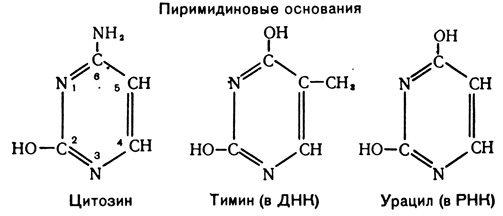
Кроме фосфата и углеводного компонента, в состав каждого нуклеотида входят по одному из пяти азотистых оснований: аденин, гуанин, цитозин и тимин (в ДНК) или урацил (в РНК). Аденин и гуанин являются производными пурина и называются поэтому пуриновыми основаниями: цитозин, урацил и тимин - производные пиримидина - носят имя пиримидиновых оснований.
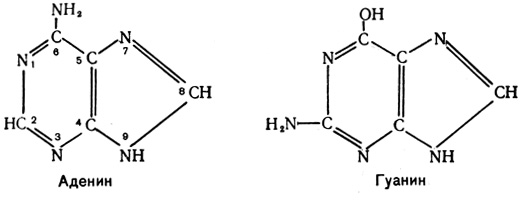
Пуриновые основания
Нуклеотиды (мононуклеотиды) являются мономерами, из которых построены молекулы нуклеиновых кислот (полинуклеотиды). В полинуклеотиде мононуклеотиды соединены между собой через остаток фосфорной кислоты, образующей эфир с 3-м углеродным атомом углеводного остатка одного нуклеотида и 5-м углеродным атомом соседнего нуклеотида. Третий атом углеводного остатка этого последнего нуклеотида соединен через другой остаток фосфорной кислоты с 5-м атомом углевода следующего нуклеотида и т. д.
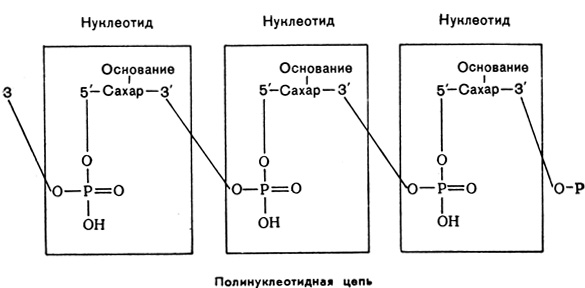
Полинуклеотидные цепи нуклеиновых кислот содержат от нескольких десятков до многих тысяч и миллионов нуклеотидных остатков. Поскольку в каждый нуклеотидный остаток входит какое-либо из азотистых оснований, то последовательность этих оснований и соответственно, нуклеотидов может быть самой различной, причем число индивидуальных последовательностей в силу наличия четырех разных оснований и большой длины цепей практически бесконечно. Таким образом, нуклеиновые кислоты представляют собой линейные не ветвящиеся полимеры нуклеотидов.
Обычно нуклеиновые основания и соответственно нуклеотиды обозначают первыми буквами: А - аденин, Г - гуанин, Ц - цитозин, У - урацил, Т - тимин. Направление цепи условно принимают от 5'-конца к 3'-концу, т. е. от замещенного гидроксила у 5-го атома углеводного остатка к гидроксилу у 3-го атома углеводного остатка (обычно незамещенному). Фосфатные остатки (иногда ими пренебрегают) обычно обозначают строчной буквой ф. Так, например, обозначение олигорибонуклеотида фГфАфГфЦфУфУфА говорит о том, что указанные нуклеотиды образуют цепь, направленную от замещенного 5'углерода гуанилового нуклеотида к конечному 3' - незамещенному остатку аденилового нуклеотида. Эту последовательность оснований в полинуклеотидной цепи называют первичной структурой нуклеиновой кислоты.
Рибонуклеиновые кислоты (РНК), как правило, состоят из одной полинуклеотидной цепочки и обладают сравнительно небольшой вязкостью в растворе. Дезоксирибонуклеиновые кислоты (ДНК) дают очень вязкие растворы, причем вязкость их зависит от скорости тока раствора. Эти свойства, а также двойное лучепреломление ДНК в потоке указывают на то, что молекулы ДНК имеют форму длинных жестких палочек.
Структура молекулы ДНК была предложена в 1953 г. молодыми учеными Джемсом Уотсоном и Френсисом Криком и получила подтверждение в дальнейших исследованиях.
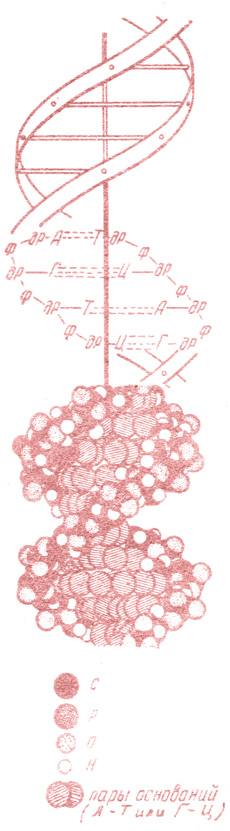
Рис. 2. Схема строения части молекулы ДНК. Атомы: С - углерод; Р - фосфор; О - кислород; Я - водород; ф - фосфат; дР - цезоксирибоза. Азотистые основания: А - аденин; Г - тимин; Г - гуанин; Ц - цитозин
Согласно этой модели (рис. 2), каждая молекула ДНК состоит из двух длинных, спирально закрученных вокруг общей оси (подобно винтовой лестнице) полинуклеотидных цепей.
Эти цепи антипараллельны (направлены в противоположные стороны), т. е. если одна цепь направлена сверху вниз (от 5' к 3' углеродным атомам), то другая цепь направлена снизу вверх. Азотистые основания, присоединенные к первому углеродному атому каждого углеродного остатка, расположены плоскостью своих колец перпендикулярно к продольной оси молекулы. Расстояние между двумя соседними основаниями составляет 0,34 нм*. Десять нуклеотидных пар образуют полный виток двойной спирали длиной 3,4 нм.
* (Нм - нанометр - единица длины, равная 1*10-9 м.)
Важнейшим условием модели Уотсона и Крика является комплементарность оснований двойной спирали ДНК. Это значит, что против каждого данного основания может находиться только комплементарное (т. е. соответствующее ему) азотистое основание. Такими комплементарными парами являются аденин и тимин и гуанин и цитозин (или коротко пары А - Т и Г - Ц).
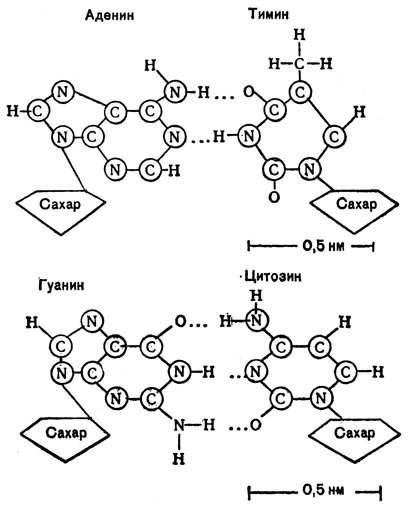
Между собой они связаны через атомы водорода так называемыми водородными связями. Между аденином и тимином таких связей две, между гуанином и цитозином - три. Каждая водородная связь очень слабая, однако три связи между гуанином и цитозином прочнее соединяют эту пару оснований, чем две связи между аденином и тимином.
Большое число водородных связей по всей длине двойной спирали довольно прочно связывает обе цепи между собой.
Две антипараллельные полинуклеотидные цепи каждой молекулы ДНК являются на всем протяжении комплементарными друг к другу, т. е. данной нуклеотидной последовательности одной цепи соответствует определенная комплементарная последовательность другой цепи Благодаря этому обстоятельству нуклеотидный состав ДНК обнаруживает определенные закономерности, получившие название правил Чаргаффа (по имени американского ученого Э. Чаргаффа, открывшего их).
- Сумма молекул пуриновых оснований равна сумме молекул пиримидиновых оснований, т. е.
А + Г = Т + Ц.
- Число молекул аденина равно числу молекул тимина, т. е.
А = Т.
- Число молекул гуанина равно числу молекул цитозина, т. е.
Г = Ц.
- Число молекул оснований с 6-аминогруппами равно числу молекул оснований с 6-оксигруппами, т. е.
А + Ц = Г + Т.
Из этого следует, что единственной вариабельной величиной в нуклеотидном составе ДНК является отношение А+Т/Г+Ц, называемое коэффициентом специфичности. Этот коэффициент колеблется в очень широких пределах у бактерий, низших растений и животных, однако у всех высших растений и у позвоночных животных он близок к 1,5. Эта последовательность нуклеотидов в полинуклеотидной цепи уникальна для каждой индивидуальной нуклеиновой кислоты. Именно эта последовательность в ДНК содержит запись всех наследственных свойств данного организма.
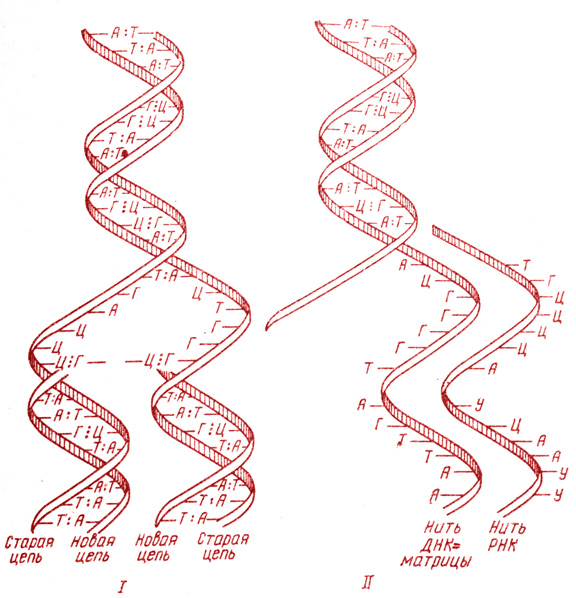
Рис. 3. Схема репликации молекулы ДНК (I) и транскрипции молекулы РНК на ДНК-матрице (II)
В S-периоде митотического цикла (см. главу II, § 5) происходит процесс удвоения количества ДНК. Этот процесс получил название репликации ДНК (рис. 3, I; рис. 4) и заключается в точном воспроизведении обеих комплементарных полинуклеотидных цепей на каждой цепи двойной спирали материнской молекулы. Репликация происходит полуконсервативным, или матричным, способом, т. е. каждая дочерняя молекула ДНК состоит из одной старой и одной вновь синтезированной, комплементарной к ней полинуклеотидной цепи. Благодаря сложному, высокоспециализированному механизму репликации две дочерние молекулы точно воспроизводят исходную молекулу ДНК, что обеспечивает упоминавшийся во введении удивительный консерватизм наследственности.
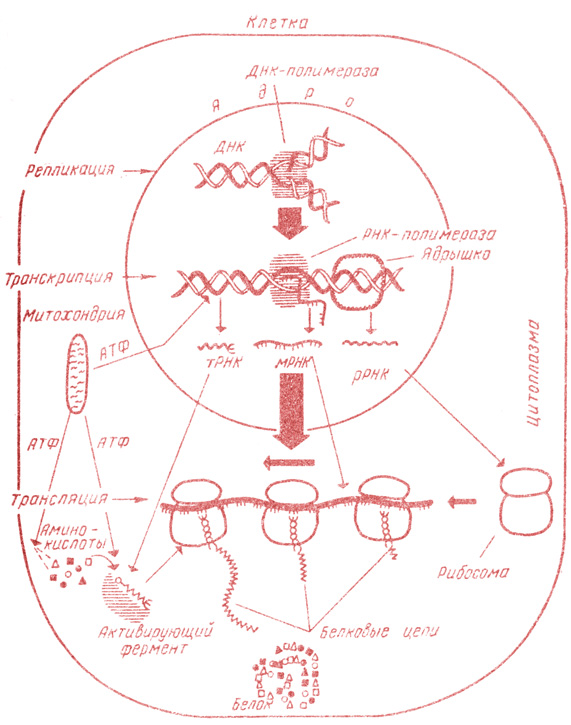
Рис. 4. Схема биосинтеза белка
Все типы РНК синтезируются на матрице ДНК (см. рис. 3, II; рис. 4) и воспроизводят последовательность нуклеотидов. При этом последовательность нуклеотидов в ДНК как бы переписывается в комплементарную последовательность нуклеотидов РНК, вследствие чего весь процесс получил название транскрипции:
Нуклеотидная последовательность РНК (место тимина в ней занимает урацил) определяет последовательность включения аминокислот в полипептидные цепи белков в процессе биосинтеза белков (трансляция) и представляет собой нуклеотидный или аминокислотный код (табл. 1).
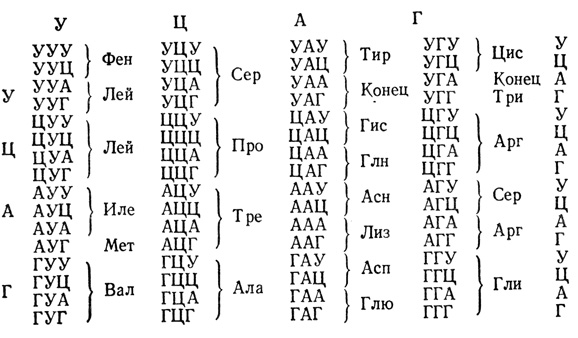
Таблица 1. Аминокислотный (нуклеотидный) код мРНК (или иРНК)
Азотистые основания: У - урацил; Ц - цитозин; А - аденин; Г - гуанин.
Аминокислоты: Ала - аланин; Арг - аргинин; Асн - аспарагин; Асп - аспарагиновая кислота; Цис - цистеин; Глн - глютамин; Глю - глютаминовая кислота; Гли - глицин; Гис - гистидин; Иле - изолейцин; Лей - лейцин; Лиз - лизин; Мет - метионин; Фен - фенилаланин; Про - пролин; Сер - серин; Тре - треонин; Три - триптофан; Тир - тирозин; Вал - валин; Конец - окончание синтеза полипептидной белковой цепи (точка терминации).
Однако не все типы РНК передают свою информацию на белок.
Различают несколько типов РНК. Главная масса РНК клетки (70-80%) приходится на рибосомную РНК (рРНК), входящую в состав рибосом. Рибосомная РНК представлена тремя видами: высокополимерной рРНК большой субъединицы с молекулярной массой 1,0-1,5*106, высокополнмерной рРНК малой субъединицы с молекулярной массой 0,5-0,8*106 и низкомолекулярной рРНК с молекулярной массой около 4*104, входящей в состав большой субъединицы. Второй тип РНК называется транспортной РНК (тРНК) и составляет 10-15% от всей РНК клетки. Она обладает относительно небольшой молекулярной массой (20-30*103) и осуществляет "доставку" аминокислот к рибосомам. Третий вид - информационная (иРНК), или матричная (мРНК),- переносит генетическую информацию от ДНК к рибосомам (где является матрицей при синтезе белковых молекул). По сравнению с двумя другими типами РНК ее содержание в клетке невелико (всего 1-10% от общего количества РНК).
Однако именно эта мРНК несет в себе аминокислотный код.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'