§ 5. Т. Г. Морган и его хромосомная теория
Томас Гент Морган родился в 1866 г., в штате Кентукки (США). Окончив в двадцать лет университет, в двадцать четыре года Морган удостаивается звания доктора наук, а в двадцать пять лет становится профессором.
С 1890 г. Морган занимается экспериментальной эмбриологией. В первом десятилетии 20-го века увлекается вопросами наследственности.

Т. Г. Морган (1866-1945)
Звучит парадоксально, но Морган вначале своей деятельности был ярым противником учения Менделя и собирался опровергать его законы на животных объектах - кроликах. Однако попечители Колумбийского университета сочли этот опыт слишком дорогостоящим. Так Морган начал свои исследования на более дешевом объекте - плодовой мушке дрозофиле и затем не только не пришел к отрицанию законов Менделя, но и стал достойным продолжателем его учения.

Н. К. Кольцов (1872-1940)
Исследователь в опытах с дрозофилой создает хромосомную теорию наследственности - крупнейшее открытие, занимающее, по выражению Н. К. Кольцова, "то же место в биологии, как молекулярная теория в химии и теория атомных структур в физике".
В 1909-1911 гг. Морган и его не менее прославленные ученики А. Стёртевант, Г. Меллер, К. Бриджес показали, что третий закон Менделя требует внесения существенных дополнений: наследственные задатки не всегда наследуются независимо; порой они передаются целыми группами - сцепленно друг с другом. Такие группы, расположенные в соответствующей хромосоме, могут перемещаться в другую гомологичную при конъюгации хромосом во время мейоза (профаза I).
Полностью хромосомная теория была сформулирована Т. Г. Морганом в период с 1911 по 1926 г. Своим появлением и дальнейшим развитием эта теория обязана не только Моргану и его школе, но и работам значительного числа ученых, как зарубежных, так и отечественных, среди которых в первую очередь следует назвать Н. К. Кольцова и А. С. Серебровского (1872-1940).
Согласно хромосомной теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенном локусе (от лат. locus - место), лежат гены. Поскольку хромосомы парны, то каждому гену одной хромосомы соответствует парный ген другой хромосомы (гомолога), лежащий в том же локусе. Эти гены могут быть одинаковыми (у гомозигот) или разным (у гетерозигот). Различные формы генов, возникающие путем мутации из исходного, называются аллелями, или аллеломорфами (от греч. алло - разный, морфа - форма). Аллели по-разному влияют на проявление признака. Если ген существует более чем в двух аллельных состояниях, то такие аллели в популяции* образуют серию так называемых множественных аллелей. Каждая особь в популяции может содержать в своем генотипе любые два (но не более) аллеля, а каждая гамета - соответственно лишь один аллель. В то же время в популяции могут находиться индивидуумы с любыми аллелями этой серии. Примером множественных аллелей могут служить аллели гемоглобина (см. главу I, § 5).
* (Популяцией (от лат. popularus - население) называется группа особей одного вида, объединенных взаимным скрещиванием, в той или иной мере изолированная от других групп особей данного вида.)
Степень доминирования в сериях аллелей может возрастать от крайнего рецессивного гена до крайнего доминантного. Можно привести большое число примеров такого типа. Так, у кроликов рецессивным геном серии множественных аллелей является ген с, обусловливающий развитие альбинизма*. Доминантным по отношению к этому гену будет ген ch гималайской (горностаевой) окраски (розовые глаза, белое тело, темные кончики носа, ушей, хвоста и конечностей); над этим геном, а также над геном с доминирует ген светло-серой окраски (шиншилловой) cch. Еще более доминантной ступенью оказывается ген агути - са (доминирует над генами с, ch и cch). Самый доминантный из всей серии ген черной окраски С доминирует над всеми "нижними ступенями аллелей" - генами c, ch, cch, са.
* (Отсутствие пигмента (см. главу VII, § 5).)
Доминантность, как и рецессивность аллелей,- не абсолютное, а относительное их свойство. Степени доминантности и рецессивности могут быть различны. Один и тот же признак может наследоваться по доминантному или рецессивному типу.
Так, например, складка над внутренним углом глаза (эпикантус) у монголоидов наследуется доминантно, а у негроидов (бушмены, готтентоты) - рецессивно.
Как правило, заново возникающие аллели рецессивны, наоборот, аллели старых сортов растений или пород животных (еще в большей степени диких видов) - доминантны.
Каждая пара хромосом характеризуется определенным набором генов, составляющих группу сцепления. Именно поэтому группы разных признаков иногда наследуются совместно друг с другом.
Так как соматические клетки дрозофилы содержат четыре пары хромосом (2n = 8), а половые - вдвое меньше (1n = 4), то у плодовой мушки насчитывается четыре группы сцепления; аналогично этому у человека число групп сцепления равно числу хромосом гаплоидного набора (23).
Для ряда организмов (дрозофила, кукуруза) и некоторых хромосом человека* составлены хромосомные, или генетические, карты, представляющие собой схематичное расположение генов в хромосомах.
* (К настоящему времени установить точную локализацию генов человека (если принять во внимание общее число генов) удалось лишь в отдельных и относительно редких случаях, например для признаков, сцепленных с половыми хромосомами.)
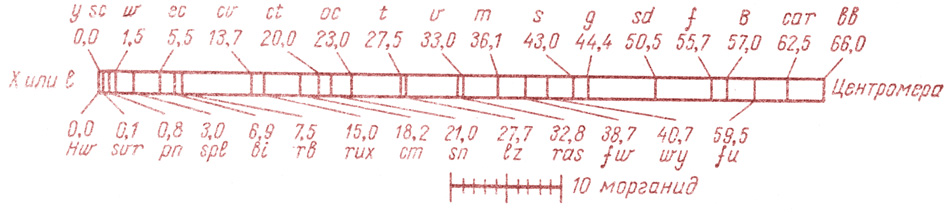
Рис. 24. Часть хромосомной карты Х-хромосомы дрозофилы (Drosophila melanogaster), составленная по данным перекреста. y (yellow) — желтое тело; sc (scute) — отсутствие некоторых щетинок; pn (prune) — черносливные (темно-коричнево-красные) глаза; w (white) — белые глаза; rb(ruby) — рубиновые глаза; cm (carmin) — карминовые глаза; ct (cut) — обрезанный край крыла; v (vermitfon) — киноварные глаза; m (miniature) — миниатюрные крылья; В (Bar) — полосковидные глаза; f (forked) — вильчатые шетинки; car (carnation) — алые глаза; bb (bobbed) — подстриженные, короткие щетинки и другие гены
В качестве примера приведем хромосомную карту части Х-хромосомы дрозофилы (рис. 24). С большей или меньшей точностью в этой карте отражены последовательность генов и расстояние между ними. Определить расстояние между генами удалось при помощи генетических и цитологических анализов кроссинговера, который происходит при конъюгации гомологичных хромосом во время зигонемы профазы I мейоза (см. главу II, § 7).
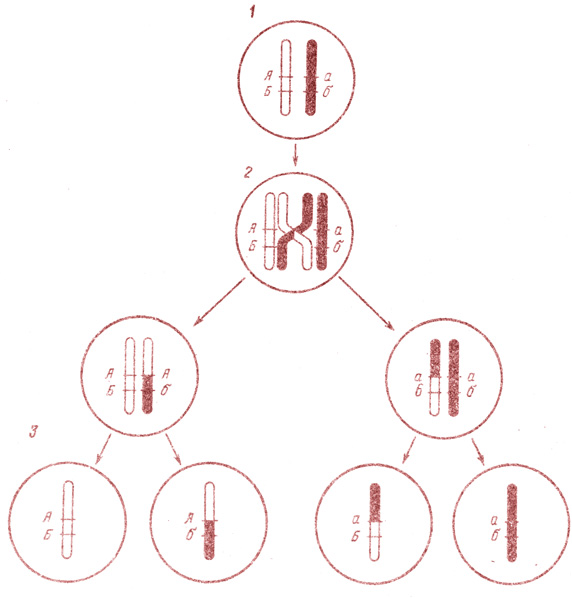
Рис. 25. Кроссинговер (схема). 1 — две гомологичные хромосомы; 2 — их перекрест во время конъюгации; 3 — две новые комбинации хромосом
Перемещение генов из одной хромосомы в другую происходит с определенной частотой, которая обратно пропорциональна расстоянию между генами: чем меньше расстояние, тем выше процент перекреста (единица расстояния между генами названа в честь Моргана морганидой и равна минимальному расстоянию в хромосоме, которое может быть измерено путем кроссинговера). Кроссинговер изображен на рис. 25.
В настоящее время известно тесное сцепление некоторых локусов генов и для них вычислен процент пере-креста. Сцепленные гены определяют, например, проявление резус-фактора и генов MN-системы крови (о наследовании свойств крови см. главу VII, § 3). В отдельных семьях удалось проследить сцепление резус-фактора с овалоцитозом (наличие примерно 80-90% эритроцитов овальной формы - аномалия протекает, как правило, без клинических проявлений), которые дают около 3% перекреста. До 9% кроссинговера наблюдается между генами, контролирующими проявления групп АВО крови и фактором Lu. Известно, что ген, влияющий на аномалию строения ногтей и колена, также сцеплен с локусами АВО-системы; процент перекреста между ними около 10. Значительно лучше изучены группы сцепления (а следовательно, и хромосомные карты) Х- и Y-хромосом человека (см. главу VII, § 6). Известно, например, что тесно связаны между собой гены, определяющие развитие дальтонизма (цветовой слепоты) и гемофилии (кровоточивости); процент перекреста между ними равен 10.
Правильность гипотезы Моргана была подтверждена в начале века Куртом Штерном (цитологические исследования) и сотрудниками Моргана Теофилусом Пайнтером (цитологом) и Кальвином Бриджесом (генетиком) на гигантских хромосомах слюнных желез личинок дрозофилы (подобных гигантским хромосомам других двукрылых). На рис. 26 показана часть гигантской хромосомы слюнной железы личинки хирономуса (мотыля).
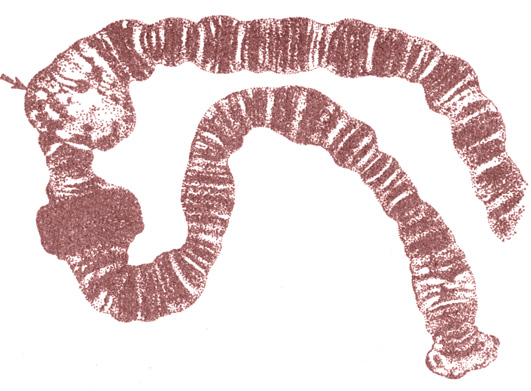
Рис. 26. Часть гигантской полинемной хромосомы слюнной железы мотыля (пуфф отмечен стрелкой)
При изучении гигантских хромосом с помощью обычного светового микроскопа хорошо заметна поперечная исчерченность, образованная чередованием светлых и более темных полос дисков - хромомеров; они образованы сильно спирализованными, плотно лежащими рядом друг с другом участками.
Формирование таких гигантских хромосом называется политенией, т. е. редупликацией хромосом без увеличения их числа. При этом редуплицированные хроматиды остаются рядом, плотно прилегая друг к другу.
Если хромосома, состоящая из пары хроматид, будет девять раз последовательно удваиваться, то число нитей (хромонем) в такой политенной хромосоме будет 1024. Благодаря частичной деспирализации хромонем длина такой хромосомы увеличивается по сравнению с обычной в 150-200 раз.
В 1925 г. Стертевант показал наличие неравного кроссинговера: в одной из гомологичных хромосом может оказаться два одинаковых локуса, в которых располагаются, например, гены, влияющие на форму глаза дрозофилы - Ваr, а в другой - ни одного локуса. Так получились мухи с резко выраженным признаком узких полосковидных глаз (ген Ultra Bar) (см. рис. 31).
Кроме цитологических доказательств правильности хромосомной теории, были проделаны генетические эксперименты - скрещивание разных рас дрозофилы. Так, среди множества сцепленных генов в плодовой мушке имеются два рецессивных гена: ген черной окраски тела (bleack) и ген зачаточных крыльев (vestigial).
Назовем их условно генами а и б. Им соответствуют два доминантных аллеля: ген серого тела и нормально развитых крыльев (А и Б). При скрещивании чистолинейных мух аабб и ААББ все первое поколение гибридов будет иметь генотип АаБб. Рассуждая теоретически, во втором поколении (F2) следует ожидать следующих результатов.
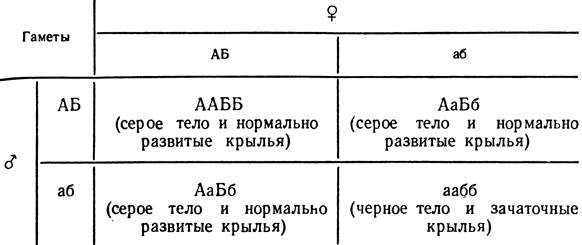
Однако в небольшом, но постоянном проценте случаев встречались необычные потомки из необычных гамет. Таких гамет в каждом скрещивании наблюдалось около 18% (9% Аб и 9% аБ).

Появление таких исключений хорошо объясняется процессом кроссинговера. Таким образом, и генетические исследования позволили установить, что нарушение сцепления - кроссинговер, приводящий к увеличению изменчивости форм, статистически постоянен.
В заключение отметим, что целый ряд положений классической генетики на сегодняшний день претерпел ряд изменений.
Мы многократно употребляли термины "доминантные" и "рецессивные" гены (аллели) и признаки. Однако исследования последних лет показали, что так называемые рецессивные гены могут фактически полностью ими не быть. Правильнее сказать, что рецессивные гены дают очень слабое видимое или невидимое проявление в фенотипе. Но и в последнем случае рецессивные аллели, внешне незаметны в фенотипе, могут быть обнаружены при помощи специальных биохимических методик. Кроме этого, один и тот же ген при одних условиях среды может вести себя как доминантный, при других - как рецессивный.
Так как развитие всех организмов происходит в зависимости и под воздействием внешней среды, то и на проявление генотипа в определенном фенотипе влияют факторы среды (температура, пища, влажность и газовый состав атмосферы, ее давление, наличие патогенных для данного организма форм, химический состав воды, почвы и пр., а для человека и явления социального порядка). В фенотипе никогда не проявляются все генотипические возможности. Поэтому в разных условиях фенотипические проявления близких генотипов могут сильно отличаться друг от друга. Таким образом, в проявлении признака участвуют (в большей или меньшей степени) как генотип, так и среда.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'