Химия мутаций
Современная теория мутагенеза стоит на "двух китах". Первый - доказательство того, что ДНК вещество, несущее наследственность. Второй - знание физико-химической структуры ДНК: модели Уотсона - Крика и механизма репликации ДНК.
Мы только что совершили путешествие в лабиринты живой клетки. Помните? ДНК-лестница раскручивается, превращаясь в гребенки. Они служат матрицей для синтеза новой цепи ДНК. Каждый зуб в них - одно из четырех оснований - притягивает к себе нуклеотид с единственно возможным (парным зубу) основанием и соединяется с ним водородными связями. Затем блоки замыкаются, и из гребенки образуется лестница. В этом процессе аденин спаривается с тимином, а гуанин - с цитозином. Но вот всегда ли? Почти всегда... С этого "почти" мы и перешагнем в теорию мутагенеза. "Законное" спаривание оснований происходит в том случае, если они находятся в совершенно определенной таутомерной форме. Биохимик скажет вам, что эта форма возникает в результате перемещения атомов водорода в молекуле азотистого основания и встречается у каждого основания. Так, например, у нормального тимина атом водорода помещается у азота в первом положении. При переходе тимина в редкую энольную форму водород перемещается к кислороду, соединенному с углеродом в шестам положении. В таутомерной форме основание спаривается "незаконно". Тимин получает возможность спариться не с аденином двойной связью, как обычно (А-Б), а тройной связью с гуанином (В-Г). При последующей репликации ДНК гуанин соединится с цитозином, который, в свою очередь, окажется на месте, где в норме должен быть тимин. Таким образом, в гене одной из нитей ДНК вместо пары А-Т окажется пара Г-Ц. Так возникнет спонтанная точечная мутация, затронувшая только одну пару оснований.
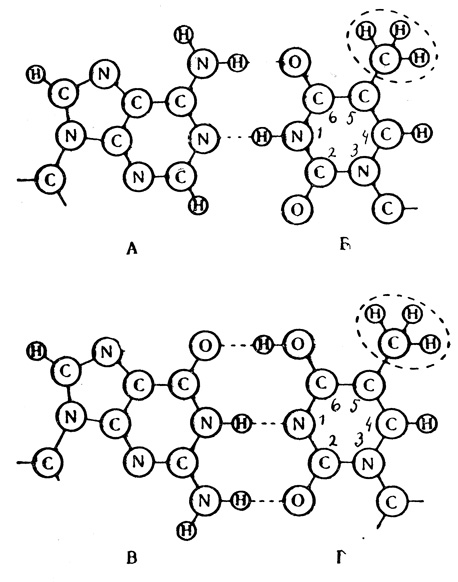
Законное (АБ) и незаконное (ВГ) спаривание тимина. А - нормальный аденин, Б - нормальный тимин, В - нормальный гуанин, Г - тимин в редкой, энольной форме
Если это место в ДНК - ген, на обязанности которого лежит контроль за выработкой какого-либо белка, точечная мутация не сильно исказит информацию. Белок будет строиться нормально. Точнее, почти нормально, ибо одна аминокислота в нем может включиться неправильно. Отразится ли это на функции белка? Все зависит от того, в каком месте произошла ошибка. Некоторые точечные мутации приводят к тому, что ген синтезирует совершенно неактивный белок. Например, известно, что замена в белке-гемоглобине всего только одной аминокислоты полностью лишает его способности быть переносчиком кислорода, то есть последствия для клетки прямо-таки катастрофические. Но если точечная мутация произошла в ином месте, белок может получиться вполне нормальным, его функция не пострадает.
К счастью, таутомерные формы оснований - редкость в мире молекул и сами спонтанные мутации - тоже: одна мутация на миллион - сто тысяч нормальных клеток. Как правило, процесс репликации ДНК в клетке идет без "ошибок".
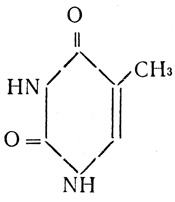
тимин
Но ведь мутации могут быть вызваны физическими и химическими агентами. И те и другие во много раз повышают их частоту. Тщательное изучение индуцированных мутаций помогло генетикам шире раскрыть двери в тайники молекулярных причин мутирования. Сделать это помог 5-бромурацил. Это вещество - близкий родственник тимина.
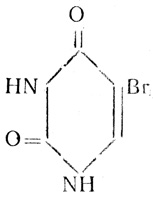
5-бромурацил
5-бромурацил отличается от тимина тем, что в бензольном кольце к углероду в пятом положении присоединена не метальная группа (CH3), а бром (Br). Поскольку при репликации ДНК тимин присоединяется к аденину в определенном месте, то эту разницу аденин просто "не заметит" и присоединит 5-бромурацил вместо тимина.
Но это не значит, что такое присоединение 5-бромурацила в цепь ДНК дает хмутацию, ведь при последующей репликации ДНК бромурацил вновь присоединит аденин, и ошибка исправится уже во втором поколении.
Опасность вовсе не в этом. Вернее, не при таком способе включения. Бромурацил повышает частоту мутаций, потому что он гораздо чаще, чем тимин, находится в таутомерной (энольной) форме. А ведь эта форма спаривается не с аденином, а с гуанином. Тогда ошибка незаконного спаривания даст точечную мутацию, которая проследует через много поколений (если, конечно, мутация не помешает существу производить себе подобных).
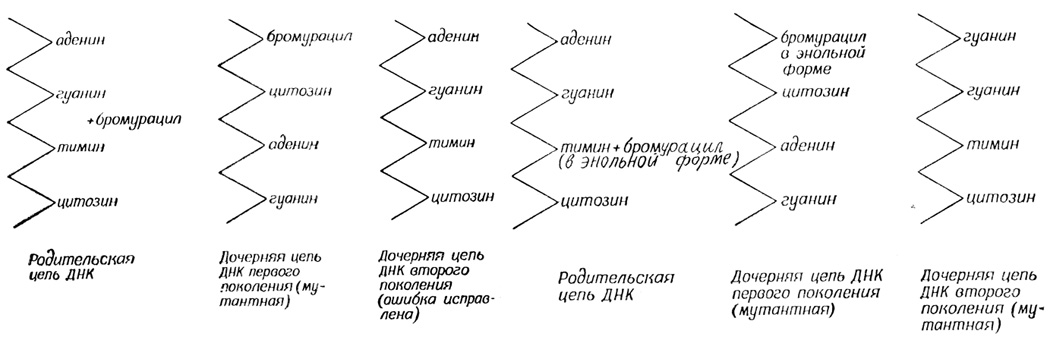
Опыты показали, что мутагенное действие бромурацила действительно осуществляется этими двумя способами. Позднее подобный механизм мутагенного действия был описан для аналога аденина - 2-аминопурина.
Но еще большую уверенность почувствовали генетики, когда нашли вещества, вызывающие мутации не только во время репликации, но и в покоящейся ДНК, например у свободного внеклеточного фага. Это оказались знакомая всем азотистая кислота и алкилирующие агенты - этилэтансульфонат и этил-метансульфонат (сокращенно их окрестили ЭЭС и ЭМС). Азотистая кислота давала окислительное дезаминирование оснований. Так, например, она переводила цитозин в урацил. Там, где появился урацил, при репликации ДНК к нему присоединится уже не гуанин, а аденин. Произойдет мутация. Тимин и урацил не содержат аминогруппы, потому им не страшна азотистая кислота, они не изменяются под ее воздействием. Вот почему принято думать, что мутации, вызванные азотистой кислотой, объясняются ее действием только на аденин и цитозин.
ЭЭС и ЭМС разрывают связь между основанием и дезоксирибозой. Можете себе представить, что за этим следует? Аденин (или иное основание) попросту уходит в цитоплазму. Гребенка теряет зуб. В образовавшуюся брешь может включиться любое основание. Если попадает верное - мутации нет, если неверное - она возникает.
Но особенно интересный сюрприз приподнесли ученым красители акридиновой серии (профлавин и другие). Они были мутагенны только в отношении бактериофага и по неизвестной до сих пор причине не действовали на бактерии. Молекула акридина, видимо, внедряется между соседними парами оснований ДНК, раздвигая их в обе стороны, так что остов ДНК несколько раскручивается. Это было показано при рентгеноструктурном анализе акридиновых мутантов фага Т4. Акридин "сидит" на ДНК, но не спаривается ни с одним из оснований. Вот почему место, где внедрился акридин, - это брешь в молекуле ДНК размером в несколько ангстрем. При репликации ДНК в новой нити брешь может заполняться любым основанием. И какое бы основание ни включилось в дочернюю реплику, оно будет лишнее. А если молекула вклинится между основаниями строящейся новой гребенки? Тогда она раздвинет ее на несколько ангстрем и окажется как раз напротив одного из оснований цепи-матрицы, к примеру, аденина. Этим она замаскирует его, да так, что комплементарный аденину партнер - тимин не сможет спариться с ним, а значит, не попадет в новую цепь и в ней недостанет одной молекулы тимина. При последующей репликации новая цепь навсегда сохранит свой дефект (нехватку тимина) - произойдет мутация типа нехватки.
Итак, мутация - устойчивое изменение информации в ДНК. Это может быть либо ошибка при репликации ДНК (как в случае присоединения таутомерных форм оснований или их аналогов), либо замена одной пары оснований другой (как в случае действия ЭЭС или ЭМС), либо вставка или выпадение одного из оснований (как в случае с акридином).
Все описанные мутации - точечные. Они касаются изменений в одном нуклеотиде ДНК. Но, как известно, мутация может проявляться в полной потере пары или нескольких пар оснований (деления), в повороте на 180 градусов блока из нескольких пар оснований (инверсия) и других внутригенных перестройках. Их называют еще хромосомные абберации. Правда, эти мутации у микробов - чрезвычайная редкость и изучены они еще мало, исключая делеции. А редкость потому, что выпадение целого участка ДНК в таком организме, как бактерия или фаг, чаще всего ведет к их гибели.
Но, кроме мутаций, в природе существует второй путь изменчивости признаков организма. Это процесс рекомбинации. Он непосредственно связан с полом и является результатом замены участка хромосомы одной особи аналогичным участком хромосомы другой особи, генетически отличающейся от первой. (Генетические отличия хромосом, как мы уже знаем, - это результат мутаций.)
Процесс рекомбинации наследственного вещества дает природе необычайно богатые возможности перегруппировки генов, что нередко нивелирует вредные мутационные изменения (ибо нормальные гены, как правило, доминируют над мутантными), приводит к наиболее выгодным сочетаниям признаков.
Изучение рекомбинаций - главная проблема генетики и вместе с тем основной метод, которым пользуются при построении генетических карт. Вот почему особенно парадоксально, что механизм этого явления до сих пор почти не изучен. Даже новейшие достижения молекулярной биологии не смогли до конца вскрыть его.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'