Опыты, опыты...
Почти четверть века назад Дарлингтон разработал свою теорию обмена наследственным веществом при гибридизации. Она была основана на поведении хромосом в мейозе: закручивание двух хроматид вокруг друг друга приводит к их чрезмерному натяжению, в результате они рвутся в точке наибольшего напряжения подобно тому, как рвутся пряди нитей в сильно перекрученном канате. Как только произошел разрыв, вызвавшие его силы ослабевают и концы разорванных нитей наследственного вещества воссоединяются. Но далеко не всегда нить соединяется со своей оторванной частью. И это понятно: после разрыва нити перемещаются, их положение меняется. Вот почему чаще всего оторванный кусок присоединяется к куску от другой хромосомы крест-накрест по месту разрыва - так образуются две рекомбинантные хромосомы. Этот механизм назвали разрывом и воссоединением, а сам процесс - кроссинговером.
Но разрыв и воссоединение в процессе кросеинговера - не единственное условие создания гибридных хромосом. Еще до Дарлингтона в 1931 году ученый Беллинг связал процесс рекомбинации с самим актом дубликации, удвоения хромосомы.
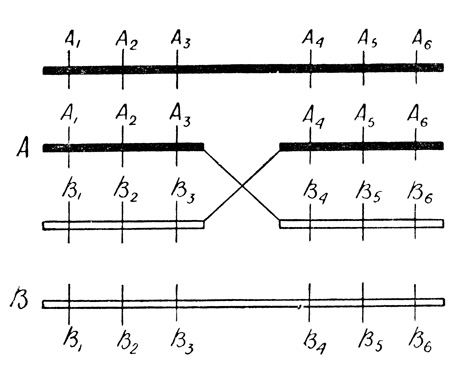
Смотрите! На следующей странице изображены две хромосомы клетки, из которой образуются половые. Одна, темная А (с признаками А1, А2 и т. д.) - от матери, вторая, светлая В (с признаками В1, В2 и т. д.) - от отца.
Но вот пришло время делиться, и хромосомы начинают строить свои копии. Одна из копий оказалась зажатой между двумя хромосомами. Возможно, во время репликации спаренных хромосом хромосома-копия, формирующаяся, скажем, вдоль материнской хромосомы (А), внезапно переходит на отцовскую (В) и продолжает строиться уже на ней, копируя ее. Когда реплика отцовской хромосомы в свою очередь дойдет до этой точки переключения, ей не остается ничего другого, как поступить по примеру первой реплики: переключиться на материнскую и копировать ее. В результате образуются две рекомбинантные хромосомы: А1, А2, А3, В4, В5, В6 и В1, В2, В3, А4, А5, А6, совсем такие же, как если бы между хромосомами А и В произошел кроссинговер. В современном изложении гипотеза такого способа рождения гибридной хромосомы получила имя - копирование с переменой матрицы.
Разница между этими двумя моделями в том, что при разрыве и воссоединении рекомбинантные новорожденные хромосомы наследуют материал непосредственно от родительских уже построенных хромосом, а при копировании с переменой матрицы рекомбинанты-хромосомы синтезируются из нового материала, получая от обоих родителей только генетическую информацию.
Какая же гипотеза правильна? Сейчас ответить на этот вопрос еще трудно. Есть данные в пользу первой гипотезы, есть данные в пользу второй, а есть и такие, которые не согласуются ни с одной из них.
Главная трудность в том, что мы по сути дела точно не знаем, что такое хромосома высших организмов, из каких молекул и по какому чертежу "сработала" ее природа. Мы только можем увидеть поведение этих хромосом в мейозе. У невидимых же (фагов и бактерий) мы знаем, как устроена хромосома. Это - кольца двухцепочечных ДНК. Но никто еще не видел, каким образом воссоединяются хромосома бактерии-матери с кусочком хромосомы бактерии-отца. А ведь мы пытаемся растолковать тонкости процесса обмена наследственным материалом у высших и у невидимок, синтезируя то, что известно о хромосомах тех и других существ. Тут-то и рождаются всяческие неточности и неувязки.
Эти неувязки породил переход генетики на микроскопические объекты. Впрочем, видимо, так и должно было случиться - истина может оказаться сложнее любой из гипотез... Например, до недавнего времени считали, что рекомбинация возникает вследствие кроссинговера между хромосомами, спаренными последовательно точка за точкой по всей длине. А Причард в 1955-1960 годах показал, что хромосомы могут спариваться не по всей длине, а только в нескольких точках, где возник случайный контакт. Об этом говорили опыты с грибком аспергиллюс и некоторыми фагами. Причард думает, что случайным может быть только спаривание в одной какой-либо области хромосомы. В любой другой области вероятность спаривания не случайна. Она больше в участках, лежащих рядом с областью первого спаривания, и меньше на дальних. Иными словами, соединение хромосомы в какой-нибудь одной точке облегчает их контакт в соседних точках.
Итак, есть две гипотезы, объясняющие механизм рекомбинаций. Однако в последнее время укрепляется все больше гипотеза разрыва и воссоединения. И вот что удивительно. Первые экспериментальные ее подтверждения были получены на... фагах, то есть на организмах, у которых этот процесс казался невероятным и для которых, собственно говоря, и была извлечена из забвения гипотеза Беллинга. Так в 1961 году Келленбергер, Вейгль, Мезельсон, Сталь и другие ученые показали, что при генетической рекомбинации фага лямбда происходит разрыв и воссоединение - физический обмен участками рекомбинирующихся фаговых хромосом. Позднее было выяснено, что рекомбинация совершается в отсутствие синтеза ДНК, что само по себе несовместимо с гипотезой избирательного копирования.
Так исчезли главные причины, по которым гипотеза разрыва отбрасывалась, она получает все более веские доказательства. В 1963 году стало ясно, что при конъюгации также происходит физический обмен между хромосомой бактерии-реципиента и фрагментом хромосомы-донора.
А в 1965 году американцы Кларк и Маргулис "напали на след" фермента, "вшивающего" в хромосому бактерии-матери фрагмент хромосомы бактерии-отца. Они выследили работу этого фермента в остроумном опыте на кишечной палочке К12. Это была женская клетка, не способная самостоятельно вырабатывать ни одной молекулы аминокислоты лейцина. А так как это жизненно важное вещество, то клетка росла только на среде с лейцином. Ее посеяли на такую среду в чашке Петри и бархатной "щеточкой" Ледерберга перенесли отпечатки выросших колоний на газон знаменитого Hfr, того, который славился выработкой лейцина и имел невидимого жильца - умеренный фаг лямбда.
Понимаете, что получилось? В месте отпечатков оказались и отцовские и материнские клетки, причем последние в меньшинстве. Покорные великой силе природы, микробы конъюгировали. Хромосома Hfr, а значит, ее гены - тот, который ведает синтезом лейцина, и другие - передались женским клеткам.
Как вы думаете, выросли на среде без лейцина микробы-гибриды или нет? Да, выросли почти все, то есть хромосома отца прошла в клетку матери и объединилась с ней.
А это что? Это - неудачница. Ее отпечаток не стал расти на среде без лейцина, вот - еще такая же... Может быть, ей не передался кусочек хромосомы с "лейцинным" геном? Нет, этого не могло быть. Передался. Живой свидетель тому - фаг лямбда. Он проникал в микробы-неудачники, а ведь его ген идет в материнскую клетку вслед за "лейцинным" геном. Значит, кусок хромосомы с "лейцинным" геном был в материнской бактерии. Тогда в чем причина неуспеха? Может быть, не удалась рекомбинация?
Этих неудачниц взяли в работу. И в какую! В ней участвовали и радиоактивные изотопы, и ультрафиолетовые лучи, и множество реактивов.
Финиш порадовал ученых чрезвычайно. Еще бы! Удалось доказать, что клетки-неудачницы не имеют ферментов, "залечивающих" хромосомный разрыв, а в клетках, в которых свершается таинство рекомбинации, они есть. Что они собой представляют, точно еще не известно. Возможно, это особый вид полимеразы. Ученые предполагают, что в хромосоме кишечной палочки К12 есть ген, который контролирует синтез фермента-"портного". Он активирует как один из этапов рекомбинации, так и реакцию, в ходе которой "зашиваются" повреждения ДНК. На карте хромосомы бактерии Кларк и Маргулис нашли место, где расположен ген, контролирующий синтез этого фермента.
Но довольно о химизме рекомбинаций. Уже давно пора вернуться к картографам хромосом, стремящимся разгадать тайны структуры гена и биосинтеза белка.
После опытов Херши и Чейз, Кайзера и многих других ученых никто уже не сомневался, что в бактерии размножается (реплицируется) чистая фаговая ДНК. В микробной клетке у нее две обязанности-воспроизводить самое себя более чем в "ста лицах" (аутокатализ) и воспроизводить белок, чтобы одеть в него своих потомков (гетерокатализ).
С первой задачей она справляется, видимо, так же, как ДНК бактерий и всего живого. Это - способ уже хорошо знакомый нам по консервативной репликации.
Но как ДНК фага воспроизводит белковые оболочки своих потомков, никто не знал. Было очевидно, что расшифровка этой тайны - путь к разгадке способа, по которому информация, записанная на ДНК, реализуется в форме синтеза того или иного белка, то есть путь к решению загадки биосинтеза белка вообще. Этот путь "вымостили" опыты по картированию хромосомы rII области фага Т4, позволившие нанести на карту хромосомы фага... молекулы вещества. Главными участниками опытов стали фаговые мутанты Сеймура Бензера.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'